Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Широтная зональность - Биогеография морского бентоса - Биогеография бентали Мирового океана
“Разные авторы выделяют между Северным полюсом и экватором три (Зенкевич, 1948; Зёрнов, 1949), четыре (Hedgpeth, 1957; Peres, 1961; Briggs, 1974), пять (Coomans, 1962), шесть (Stephenson, 1947; Hall, 1964; Кусакин и др., 1975), семь (Зинова, 1962), девять (Семёнов, 1977) зон и подзон” (Несис, 1985: 54).
“Произвольное использование в качестве объекта изучения биогеографии наряду с ареалом условий обитания в одних случаях исказило выявленные биогеографические закономерности, в других — просто помешало их выявить. В настоящее время нет ни одной системы, отражающей зональную дифференциацию морских организмов, которая в большей или меньшей мере не грешила бы этим недостатком” (Перестенко, 1982: 100).
Широтная зональность бентали принципиально отличается от широтной зональности суши. На суше широтную зональность выделяют на основе выделения типов растительности, являющейся основой ЦС суши (см. Главу 3). Поскольку роль растительности в бентали значима лишь в верхней сублиторали, а глубже пренебрежимо мала, авторы схем широтной зональности бентали пытаются разбить ось экватор-полюс на отрезки, соответствующие наиболее крупномасштабным биогеографическим выделам. Это легко сделать вдоль одного берега океана, но подобная схема фактически является повторением обычной частной биогеографической схемы и в её существовании немного глобального смысла. Сложности начинаются при попытке свести схемы, построенные от каждого полюса к экватору вдоль каждого берега материка, в одну общую. В разных схемах широтной зональности общими являются только эти две опорные точки: экватор и полюс. Количество же широтных зон вдоль каждого берега любой биогеограф волен выбирать в зависимости от его представлений и методов, что, собственно говоря, и делают. Ещё больше возможностей представляется при проведении широтной зональности в пределах отдельного океана и ещё больше — в пределах всего Мирового океана. Поскольку нет общепринятых методов ни районирования, ни сравнения выделенных районов и их группировки, в литературе наблюдается достаточно большое разнообразие схем широтной зональности, особенно, если учесть малое число исследователей, занимающихся их построением. При этом обычная практика состоит в объединении регионов, расположенных в сходных условиях, а не населённых сходными биотами, проведение границ между зонами в местах наиболее резких градиентов абиоты, а не смены биот. “Широтная зона — результат объединения районов, выделенных по фаунистическим показателям, но объединённых по абиотическому признаку” (Миронов, 1981: 12). Примером подобной схемы может служить широко известная схема В.Г. Богорова и Л.А. Зенкевича (1966) (Рис. 5.52).
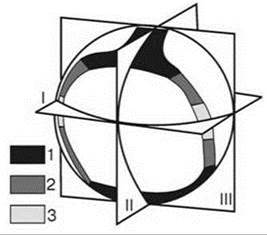
Рис. 5.52. Три плоскости симметрии Мирового океана.
Плоскости симметрии: I — экваториальная, II и III — меридиональные.
Природные зоны: 1 — полярные, 2 — умеренные, 3 — экваториальная (Богоров, Зенкевич, 1966).
Как абиотический признак часто используют температуру, например среднемесячные поверхностные изотермы самого тёплого месяца 25°C, 20°C, 15°C и 10°C для проведения границ, соответственно, тропического, субтропического, тёплого умеренного, холодного умеренного и арктического поясов. Очевидно, хорошего совпадения этих изотерм с биогеографическими границами ожидать не следует, и действительно, авторам приходится объяснять эти отклонения или использовать в каждом случае (например, для объяснения положения границ у разных берегов одного океана) разные показатели. Строго говоря, эти зоны на самом деле являются не биогеографическими, а климатическими, отражающими широтную зональность в распределении поступления солнечной энергии на Землю. Конечно, распределение жизни на Земле определяют и абиотические факторы, в частности температура, но сущность схемы определяет логическое основание: распределение абиотических факторов (в данном случае температуры) или распределение жизни.
Как отмечал К.Н. Несис (1982: 115), “сопоставление схем для противоположных сторон океана и разных океанов будет фаунистически обосновано лишь при значительном числе общих амфиокеанических видов. Число таких видов велико в приполярных областях, где преобладают циркумглобальные ареалы, и прогрессивно снижается при движении к экватору”. Изучение конкретных ареалов показывает, что они обычно ассиметричны. Примеры асимметрии в амфибореальных ареалах приведены на Рис. 5.53—5.55. Но в разорванных ареалах асимметрия может быть объяснена и тем, что в разных частях ареала обитают разные таксоны (подвиды, расы, виды). Более убедительной является асимметрия сплошных ареалов (Рис. 2.8—2.10 и Рис. 5.56—5.58). Причём эта асимметрия разная для разных видов: одни распространены шире в Атлантике, другие — в Пацифике, вдоль разных берегов одного океана виды также обычно распространены по-разному. Встречающиеся в разных океанах виды входят в состав разных широтных поясов, поскольку распространение видов обусловлено не только их биологией и абиотическими условиями, но и ареалами тех ЦС, в состав которых они входят. Число примеров асимметричных ареалов можно значительно увеличить, в реальности сложно найти примеры симметричных ареалов видов бентоса. При подборе ареалов для иллюстраций я легко нашел примеры ассиметричных ареалов и с трудом — примеры симметричных. По-видимому, симметричные ареалы — большая редкость. Справедливости ради надо отметить, что примеры симметричных ареалов пелагических видов многочисленны и именно их приводят в качестве иллюстраций симметрии амфибореальных ареалов.
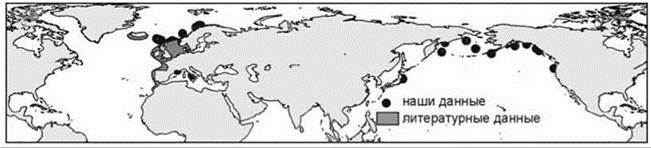
Рис. 5.53. Симметричный амфибореальный ареал: атлантическая часть = тихоокеанской полихета Pectinaira auricoma.
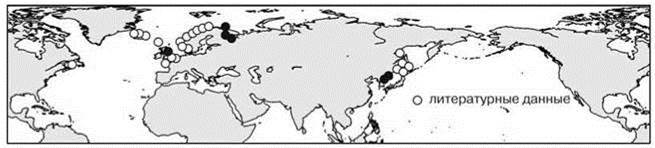
Рис. 5.54. Асимметричный амфибореальный ареал: атлантическая часть > тихоокеанской полихета Amphitrite figulus.
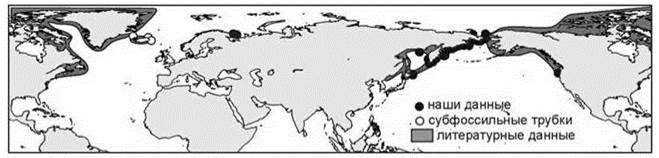
Рис. 5.55. Асимметричный амфибореальный ареал: атлантической часть < тихоокеанской полихета Pectinaria granulata.
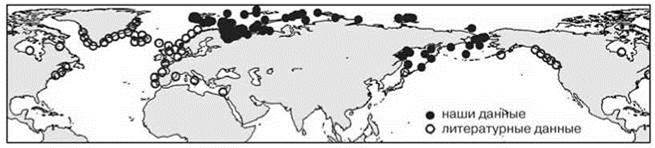
Рис. 5.56. Симметричный аркто-бореальный ареал: атлантическая часть = тихоокеанской полихета Amphitrite cirrata.
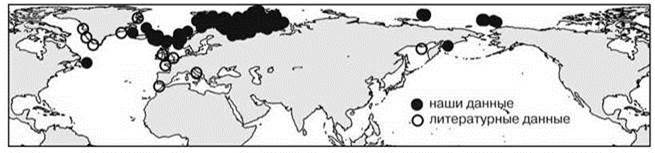
Рис. 5.57. Асимметричный аркто-бореальный ареал: атлантическая часть > тихоокеанской полихета Axionice maculata.
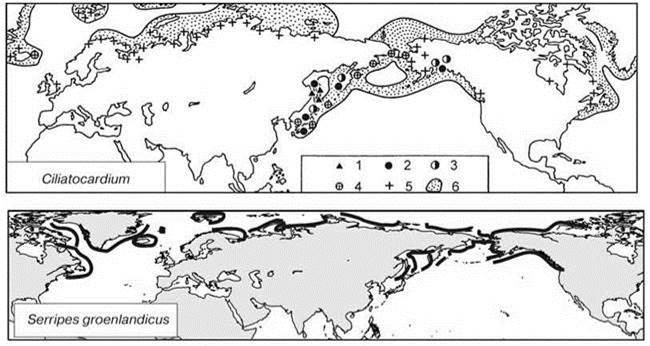
Рис. 5.58. Асимметричный аркто-бореальный ареал двустворчатых моллюсков Cardiidae: тихоокеанская часть > атлантической.
1 — эоцен, 2 — олигоцен, 3 — миоцен, 4 — плиоцен, 5 — плио-плейстоцен и плейстоцен, 6 — современный ареал. По Скарлато (1981) и Кафанову, Кудряшову (2000), распространение обоих видов исправлено по Brattegard, Holthe (1997): Ciliatocardium ciliatum встречается чуть южнее, чем указывает О.А. Скарлато, тогда как Serripes groenlandicus не заходит так далеко на юг, как показано на карте А.И. Кафанова.
* fms — сокращение от fathom — морская сажень. 1 fms равна: международная — 1,8288 м = 6 футам; английская — 1/1000 морской мили (6,080 футов) = 1,853 184 м.