Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Биполярное - Некоторые типы распространения - Схемы биогеографического районирования - Биогеография бентали Мирового океана
Названия некоторых типов ареалов часто используют в литературе, поэтому целесообразно рассмотреть их более детально.
Биполярное
Многие таксоны (виды, рода, семейства) имеют разрыв ареалов в тропиках. Такой ареал в русскойзычной литературе получил название “биполярный”. Надо отметить, что, несмотря на то, что в названии типа распространения присутствует слово “полюс”, ареалы таксонов с биполярным распространением в Северном полушарии никогда не ограничены арктическими водами, все они обитают в бореальных и/или субтропических водах, хотя могут встречаться и в арктических. Поэтому более правильно называть такой ареал так, как это принято в англоязычной литературе — “антитропический”.
Наиболее универсальна теория, связывающая явление биполярности с отдельными самостоятельными актами божественного творения (Dana, 1853; Agassiz, 1857 и Sclater, 1858). Однако в настоящее время эта теория не пользуется популярностью в научных кругах, и биполярное распространение объясняют, не привлекая помощь свыше. Не существует общего объяснения причин такого распространения, поскольку в действительности под биполярным распространением подразумевают разные типы распространения, каждый из которых имеет свою причину.
Таксоны с тропическими ареалами обычно имеют разрывы в Индо-Вест-Пацифическом треугольнике, районе с наибольшим в настоящее время видовым разнообразием, в котором и в настоящее время, по-видимому, идёт интенсивное видообразовании. Такой ареал J.C. Briggs (1995) объясняет вытеснением из этого района видов новыми. Таксономическое разнообразие также относится к хищникам, паразитам и возбудителям болезней. Каждый из этих факторов также может играть свою роль в образовании разрыва.
Таксоны, ареал которых не включает тропики, скорее всего и возникли за их пределами, поэтому биполярность в данном случае нельзя объяснить вытеснением вида из тропических районов, ибо его там никогда и не было. В случае эврибатных бореальных и эврибатных субтропических таксонов биполярность можно объяснить их способностью обитать на глубинах склона, глубже тропических вод. Фактически, они могут “подныривать” под них, делая ареал единым. Подобные единые ареалы, очевидно, были более распространенным явлением в ледниковые эпохи, когда расстояние между умеренными регионами по разные стороны экватора было меньше, особенно у восточных берегов океанов. И в настоящее время имеются виды с подобными ареалами. Хрестоматийным является распространение Eukronia hamata (Chaetognatha). Этот космополитический вид в умеренных и холодных водах обитает на небольших глубинах, а в низких широтах — на больших. Виды с таким ареалом, по мнению J.C. Briggs'a (1995) возникли преимущественно в Северной Пацифике — одном из основных центров современного видообразования за пределами тропиков, затем вселились в Северную Атлантику, а затем в Южное полушарие. В дальнейшем вид сначала исчезает в Северной Пацифике, затем в Северной Атлантике, а затем в разных частях ареала в Южном полушарии.
Распространение верхнесублиторальных бореальных таксонов представляет наибольшие сложности для интерпретации. Эти виды неспособны “подныривать” под тёплые тропические воды. В умеренных водах Северного и Южного полушарий обычно обитают те же или близкие виды, что свидетельствует о недавнем возникновении этого явления. Одним из наиболее наглядных примеров является распространение ламинариевых водорослей. Обычное объяснение — замыкание ареала в период максимального оледенения за счёт охлаждения тропиков (Берг, 1920, 1947). Эту точку зрения в отечественной литературе иногда считают “наилучшим объяснением” (Зенкевич, 1951). Однако факты не подтверждают эту гипотезу. Прежде всего, суммарное количество тепла, поступающего на Землю, скорее всего, постоянно, по крайней мере в столь малый промежуток времени. Следовательно, охлаждение в одном месте (у полюсов) должно было привести к нагреву в другом. Т. е. температура воды в тропиках должна была повыситься во время оледенений, а не понизиться, хотя может быть и не столь сильно, как должно было бы быть при абсолютном постоянстве поступающей на Землю энергии, поскольку альбедо (отражающая способность) ледников и снега гораздо больше, т. е. потери на отражение в ледниковые периоды были выше. Палеотемпературные данные не показывают заметного охлаждения в тропиках во время оледенений.
Детальный анализ распространения видов также приводит к мысли о том, что миграции таких видов через охладившиеся тропики не имели место. Дело в том, что большая часть данных по биполярному распространению относится к таксонам, обитающим в Тихом океане. Посмотрим, что наблюдается в Атлантике. После открытия 3,5 млн. лет назад Берингова пролива Северо-Тихоокеанская фауна проникла в Северную Атлантику. Оледенения развились гораздо позже этого. Однако, ни один из видов моллюсков, проникших в Атлантику из Пацифики (а их было около 300) не проник на юг Африки (Vermeij, 1992). В то же время, согласно другим данным около 20% видов Opistobranchia, обитающих на юге Африки, также обитает и у Европы (Gosliner, 1987, цит. по: Briggs, 1995). Таким образом, данные по моллюскам свидетельствуют о том, что охлаждение воды в период оледенения не может объяснить биполярное распространение мелководных бореальных видов.
Наиболее вероятной причиной недавнего возникновения биполярного распространения является миграция умеренно-тепловодных верхнесублиторальных видов по апвеллингам. Как уже говорилось ранее, в период оледенений усиливается вертикальная циркуляция Мирового океана, что приводит к усилению апвеллингов и уменьшению тропических районов между ними. Перестройка системы течений приводит также к миграции областей апвеллингов, менявшей площади, занятые тропическими районами. Апвеллинги наиболее развиты у восточных берегов (Рис. 5.9), поэтому биоты тропических районов восточных берегов океанов должны обедняться значительно сильнее по сравнению с западными. И действительно, факты свидетельствуют не о простом количественном обеднении тропических восточных районов, как следовало бы ожидать из геоморфологических данных (у западных берегов расположены архипелаги, тогда как восточные практически лишены островов, площадь шельфов у западных берегов гораздо больше). Восточные берега беднее качественно: целые типы экосистем были утрачены в периоды оледенений. Достаточно вспомнить исчезновение коралловых рифов у тихоокеанских берегов Америки и полное отсутствие морских лугов в восточных тропических районах Атлантики и Пацифики (подробнее см. Главу 1, раздел “Причины и скорость образования новых видов и экосистем”). Некоторые из них позднее восстановились, но всё же их таксономическое разнообразие осталось гораздо ниже, чем можно было бы ожидать.
Однако есть примеры биполярного распространения таксонов более высокого ранга. Очевидно, что плейстоценовое оледенение здесь не при чём. Пример этого — бесчелюстные, возникшие в ордовике. Они проникли в пресные воды в позднем силуре и раннем девоне. В настоящее время бесчелюстные представлены двумя классами — миноги и миксины. Из них миноги — пресноводные животные, некоторые из которых — проходные (т. е. нерестятся в пресной воде, а живут в морской). Распространение миног типично биполярное (Рис. 5.75). В Северном полушарии обитает одно подсемейство, в Южном — два других. Столь сильные различия свидетельствуют о достаточно долгой их изоляции, большей, чем прошло со времени последнего ледникового периода. J.A. Crame (1993) нашел, что биполярность в распространении моллюсков может быть прослежена по крайней мере до ранней юры.
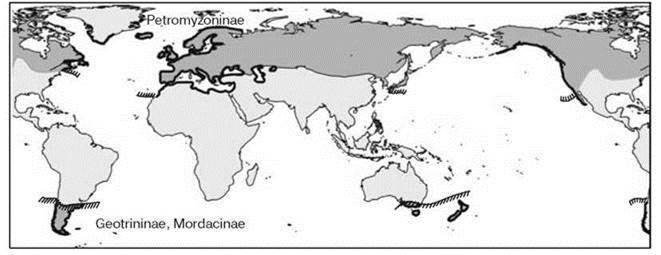
Рис. 5.75. Биполярное распространение миног.