Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Основные неогеновые центры расселения бентоса - Схемы биогеографического районирования - Биогеография бентали Мирового океана
Наиболее резко выраженные центры таксономического разнообразия расположены в районах, где ареалы ЦС наиболее велики и где через них проходит наибольший поток энергии. На шельфе таковыми являются тропические районы. По крайне мере, с середины мезозоя наиболее выраженный центр разнообразия морской биоты — море Тетис, особенно его западная часть. По мере закрывания Тетиса центр разнообразия начал смещаться восточнее, сначала, вероятно, в район, который образовался в результате сближения Индостана с Азией (где ныне располагаются горные системы от Кавказа до Гималаев). Район этот, по-видимому, был сходен с современным Индо-Малайским (включал обширные шельфы и крупный архипелаг), размеры его, вероятно, были даже больше. Постепенно площадь морей в данном районе также существенно уменьшилась, но одновременно ещё восточнее в результате сближения Австралии с Азией начал формироваться другой район, который и стал новым центром разнообразия, существующим и поныне Рис. 5.78. Он включает Индо-Малайский архипелаг и прилегающие шельфы Азии и Австралии. Этот район — центр не только разнообразия, но расселения герматипных кораллов (Stehli, Wells, 1971), иглокожих (Дьяконов, 1950; Миронов, 1983, 1985a); рыб (Allen, 1975; Mora et al., 2003) бентоса в целом (Briggs, 1974, 1984, 1992, 1999, 2000, 2003; Ekman, 1935, 1953; Myers, 1999), причём не только в тропические шельфовые районы, но и в высокие широты и на материковый склон.
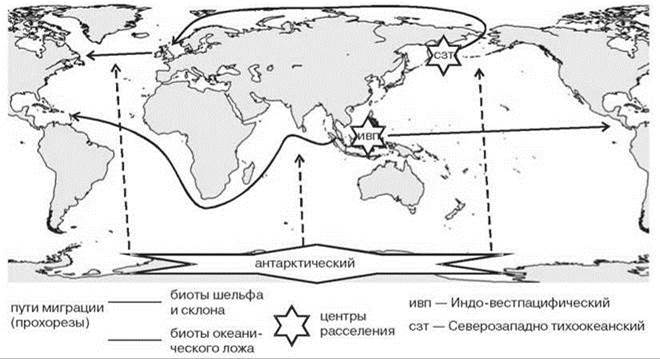
Рис. 5.78. Основные неогеновые центры расселения и направления прохорезов.
Среди всех районов по таксономическому разнообразию резко выделяется Индо-Малайский регион. Так, моллюсков там обитает 6000 видов (из 27000, известных в Мировом океане), 800 видов иглокожих (из 6000), 4000 видов рыб (из 20000). Впрочем, это не удивительно. Мелководья этого района имеют площадь 6,5 млн. км2, тогда как всех остальных тропических регионов вместе взятых — только 2 млн. км2. К тому же, эти мелководья разбиты на большое количество более или менее изолированных участков, что приводит к частичной изоляции. Колебания уровня воды в кайнозое, несомненно, изменяли степень изоляции и связи разных его участков, как за счёт возникновения и устранения преград в виде суши, так и за счёт неизбежного изменения системы течений.
G.J. Vermeij (1978) приводит многочисленные примеры родов, обитавших ранее (судя по палеонтологическим данным) циркумтропически, а ныне являющихся палеоэндемами Индо-Малайского региона. Различия других тропических регионов также в значительной степени объясняются дифференцированным вымиранием. Имеется много причин, по которым таксоны, имевшие в прошлом пантропическое распространение, ныне отсутствуют в одном или нескольких регионах или, что наблюдается часто, ограничены в своём распространении Индо-Малайским регионом:
— Климатические изменения в разных районах тропиков были неодинаковыми, Индо-Малайский регион — единственный, в котором не было существенного изменения абиотических условий с момента его возникновения. Выше уже был приведён пример восточной тропической Пацифики, в котором в недавнее время вымерли коралловые рифы, головоногие моллюски внешней части шельфа, брахиоподы и исчезли луга морских трав. Аналогичные события были и в восточной тропической Атлантике, там также сейчас отсутствуют луга морских трав и брахиоподы. Существенное вымирание было и в Вест-Индии.
— Большая площадь Индо-Малайского региона определяет то, что в разных его районах условия весьма разнообразны, причём разные биотопы занимают в его пределах существенные площади. Это позволяет существовать разнообразным специфическим ассоциациям и СС.
Сам Индо-Малайский регион также неоднороден:
— Наибольшее число видов отмечено из его центра, показанного на карте треугольником, впервые выделенного J.C. Briggs. В пределах некоторых семейств и родов к этому треугольнику приурочены наиболее специализированные формы, тогда как генералисты обитают по его периферии.
— Число видов и родов мадрепоровых рифообразующих кораллов уменьшается по мере удаления от этого треугольника, в то время как средний возраст родов, судя по палеонтологическим данным, по мере удаления от этого треугольника растёт. Растёт и возраст Balanoida и стебельчатых морских лилий.
— Имеются ареалы видов с разрывом в треугольнике Индо-Малайского региона, где такие виды замещаются другими, вероятно, более молодыми.
J.C. Briggs (1995) полагает, что эти данные свидетельствуют о непрерывно идущем процессе видообразования в этом регионе. Много весьма интересных с общебиологической точки зрения вопросов может быть изучено здесь. К сожалению, он изучен крайне недостаточно. Например, число полихет, известных из Индо-Малайского архипелага, меньше, чем известных с шельфа Северного Ледовитого океана, хотя, судя по другим, лучше изученным группам бентоса, разница должна быть на порядок. Стремительное разрушение человеком коралловых рифов Индо-Малайского архипелага — проблема мирового масштаба. Эта проблема даже более значимая, чем уничтожение тропических лесов, поскольку рифы, в отличие от лесов, не только центр разнообразия, но и играют существенную роль в связывании СО2 атмосферы.
Имеющиеся данные показывают, что некоторые из мелководных таксонов, несомненно Индо-Малайского происхождения, оказались способными преодолеть биогеографические барьеры и заселили соседние регионы.
Отнюдь не любой район со сложной геоморфологией, даже если он тропический, становится центром расселения. Например, Мексиканский залив и Карибское море (Вест-Индия) как центр разнообразия и расселения в настоящее время значительно уступает Индо-Малайскому, причём разница эта не пропорциональна разнице в площадях. По-видимому, малое значение данного района как центра расселения связано, прежде всего, с происходившими здесь эпизодами массового уменьшения числа видов. Такие события особенно хорошо документированы для плейстоцена.
В умеренных широтах Северного полушария и как центр разнообразия, и как центр расселения наиболее заметен Северозападнопацифический центр. Северо-пацифическая и аркто-атлантическая биоты имеют скорее всего одинаковый возраст. В конце палеоцена (57 млн. лет назад) произошло быстрое глобальное потепление. Поверхностная температура воды в Антарктике достигала 18°С, что привело к исчезновению всех умеренно-теплолюбивых морских биот. Вновь условия для их возникновения сформировались 25 млн. лет назад с установлением существенного температурного градиента между экватором и полюсом, поэтому северо-пацифическая и северо-атлантическая биоты не могут быть древнее 25 млн. лет. Площади, занимаемые этими биотами, были сходны. Тем не менее, в настоящее время (т. е. после совершившегося обмена) тихоокеанская шельфовая фауна вдвое богаче атлантической. Логично предположить, что различия в видовом богатстве североатлантической и северотихоокеанской биот в значительной степени обусловлены различиями в видовом богатстве донорских биот. Атланто-арктическая фауна с юга граничила с центральноамериканской и восточно-атлантической тепловодными фаунами, тогда как северотихоокеанская — с той же центрально-американской (Панамского перешейка тогда не было) и индо-малайской. Восточно-атлантическая биота уже тогда была гораздо беднее индо-малайской.
Другая причина большего видового богатства северотихоокеанской биоты — миграции нотальных биот. Поскольку охлаждение умеренных широт в Южном полушарии произошло, как минимум, на 10 млн. лет раньше, чем в Северном, нотальные биоты сформировались гораздо раньше. При наличии подходящих условий (а они были особенно благоприятны в периоды оледенений) происходил обмен бореальных и нотальных биот. Выше Главе 1 (раздел “Причины и скорость образования новых видов и экосистем”) было показано, что наиболее подходящий путь такого обмена — вдоль тихоокеанских берегов Америки. Дополнительные аргументы приведены также в разделе о биполярных ареалах.
После установления контакта между северо-аталнтической и север-пацифической биотами преимущественное направление миграций было из Пацифики в Атлантику, причины этого и детали обмена разобраны выше, в последнем разделе Главы 1.
Теоретически существует два возможных пути миграции тихоокеанской шельфовой биоты в Атлантику: вдоль Евразии и вдоль Северной Америки. Последний путь, начиная с работ S. Ekman (1935), Е.Ф. Гурьяновой (1957) и, особенно, после работы К.Н. Несиса (1961), часто считают главным или даже единственным. Эту точку зрения подкрепляют тремя основными группами аргументов: (1) географическими: путь в Атлантику через Канадский арктический архипелаг короче, чем вдоль Евразии, Е.Ф. Гурьянова даже предполагает, что хребет Ломоносова выступал над поверхностью моря, делая невозможным евразийский миграционный путь; (2) гидрологическими: поверхностные течения в период миграции бореальных видов скорее всего были теми же, что и ныне, т. е. шли из Атлантики вдоль Евразии, а от Чукотского моря вдоль Америки; при этом сибирские моря распреснены, что делает невозможным для морских видов миграцию вдоль Сибири; (3) существуют виды, обитающие в Пацифике и только в западной Атлантике, например, краб Chinoecetes opilio и плоский ёж Echinarachnius parma. Относительно второго первого аргумента можно отметить, что некоторые виды, проникшие в Атлантику из Пацифики не имеют пелагических личинок, поэтому направление течений для них несущественно. Распреснение же охватывает лишь поверхностные горизонты, тогда как среди видов, мигрировавших из Пацифики очень немногие столь стенобатны, чтобы для них была невозможной миграция под распреснёнными водами. Относительно третьего аргумента сам К.Н. Несис пишет, что в конце плиоцена “нынешние тихоокеанские-западноатлантические виды моллюсков обитали в Англии и Исландии” (1961: 895). Также имеются виды с тихоокеанско-восточноатлантическим ареалом, причём не только мелководные, как утверждает К.Н. Несис, но и обитающие в нижней сублиторали и глубже (например, полихета Pectinaria auricoma). Л.П. Перестенко (1994), исследовав современное распространение красных водорослей (т. е. видов верхнесублиторальных, на которых постулируемые К.Н. Несисом факторы должны были сказываться в наибольшей степени), пришла к прямо противоположному выводу: о преимущественно евразийском пути обмена. Среди видов исследованной ею флоры нет видов, обитающих только в западной Атлантике и западной Пацифике, хотя есть виды, обитающие только в западной Пацифике и восточной Атлантике. Л.П. Перестенко указывает, что вселение западнотихоокеанских видов происходит в европейские, а не американские воды.
G.J. Vermeij (1991) объясняет тихоокеанско-западноатлантические ареалы относительной бедностью западноатлантической биоты по сравнению с восточноатлантической. Это привело к появлению в западной Атлантике сравнительно больших свободных ресурсов, которые и стали потреблять вселившиеся из Тихого океана виды. Поэтому, вероятно, более правильно другое объяснение: тихоокеанских видов в западной Атлантике больше не потому, что им было туда легче попасть, а потому, что им там было легче закрепиться. В частности, оказалось, что краб-стригун Chionoecets opilio способен пройти пешком из Чукотского моря в Баренцево: в 1993 г. он был пойман на полпути (в море Лаптевых) на расстоянии в тысячи км от границы ближайшей части его современного ареала. Совершенно очевидно, что этот путь взрослая особь краба прошла пешком, причём по евразийскому пути. Каких-либо палеонтологических следов таких миграций, естественно, остаться не могло.
Таксоны, возникшие в северотихоокеанском центре проникли даже в Антарктику, преимущественно опускаясь в низких широтах на глубины материкового склона. Среди рыб к таким таксонам относятся семейства Zoarcidae и Liparididae, антарктическая фауна которых насчитывает 90 видов (> 40% антарктической ихтиофауны) (Briggs, 2006).
На материковом склоне наиболее ярко выражен Западнопацифический центр разнообразия и расселения (Дьяконов, 1950; Миронов, 1980, 1993; Whatley, 1987), расположенный там же, где Индо-Вест-Пацифический, но на больших глубинах. Батиальная фауна этого района расселялась и расселяется по всему Мировому океану, но, особенно, в низких широтах.
На океаническом ложе (не на шельфе!) условия для формирования повышенного таксономического разнообразия создаются там, где на дно поступает большой поток энергии (в виде детрита), т. е. где экосистемы фотического слоя имеют повышенную продукцию и плохо сбалансированы — в районах апвеллингов и полярных фронтов. Но, если в районах полярных фронтов происходит опускание поверхностных вод, из-за чего придонные слои воды содержат достаточно много кислорода, то в районах аплеллингов происходит подъём воды, и кислород, содержащийся в воде, дошедшей до района апвеллинга, которого и так часто меньше 50% от насыщения, расходуется на окисление поступающей сверху органики, поэтому условия существования бентоса там не столь благоприятны. К тому же полярные фронты представляют собой гораздо более крупномасштабные структуры. Наиболее выраженным центром разнообразия и расселения абиссальной фауны является район Антарктической конвергенции (Беляев, 1977; Гебрук, 1990; Кусакин, 1973; Миронов, 1980, 1982, 19856, 1993). Абиссальные таксоны антарктического происхождения составляют основу абиссальной фауны всего Мирового океана. Сама же абиссальная фауна Антарктики возникла преимущественно за счёт таксонов низкоширотного происхождения.