Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Типы ареалов пелагических видов - Планктон и нектон - Биогеография пелагиали Мирового океана
Обитатели пелагиали образуют две естественные группы: планктон и нектон. Организмы планктона парят в толще воды и не способны к существенным перемещениям по горизонтали, хотя многие способны к вертикальным миграциям, порой на километр и более. В зависимости от таксономической принадлежности планктон делят на вириопланктон (вирусы), бактериопланктон, фитопланктон (водоросли), зоопланктон (простейшие, беспозвоночные, за исключением некоторых головоногих) и многие костистые рыбы (ихтиопланктон). Хрящевых рыб в составе планктона нет.
Главное различие планктона и нектона — не размер, а способность к длительным активным миграциям по горизонтали у нектона и отсутствие такой способности у планктона. Большая часть пелагических костистых рыб не способна к длительным миграциям и их следует включать в состав планктона. Они входят в ту же размерную группу, что и эуфаузииды, креветки и мелкие кальмары, которую в отечественной литературе принято называть макропланктоном, в иностранной — forage organisms (кормовые организмы), поскольку они являются основной пищей промысловых видов нектона (тунцов, китов). Крупные планктонные рыбы двигаются ундулирующими движениями плавников. Самые крупные виды планктона: луна-рыба (Mola) — свыше 3 м при весе 1500 кг, сельдяные короли Regalecus — до 6 м при весе более 250 кг, пелагическая гастропода Carinaria cristata до 50 см длины. Среди крупных головоногих также есть планктонные виды (у кальмаров Thysanotheuthis spp. длина мантии до 1 м), добычу они добывают, пассивно паря в тоще воды растопырив щупальца (у некоторых щупальца снабжены светящимися органами, приманивающими добычу). Нейтральную плавучесть обеспечивают воздушные пузыри раковин или накопление хлористого аммония в вакуолях или целоме. Сходный образ жизни ведут и самые крупные головоногие — Architeuthis, достигающие 18 м длины (мантия — до 5 м), но они совершают нерестовые миграции, поэтому их следует относить к нектону.
Хотя планктон — это обитатели пелагиали, только часть видов планктона в своём существовании не связана с дном. Такие виды называют голопланктон. Виды, часть жизненного цикла которых связана с дном, называют “меропланктон”. Для мелководных (обитателей глубин шельфа) видов планктона и нектона, тесно связанных своей биологией с дном, используют термин неритические виды. Виды открытого океана называют океанический планктон. Роль бентоса в неритических сообществах часто весьма велика. Зоопланктон в значительной своей части, преимущественно на малых глубинах в периоды размножения бентоса, состоит из его личинок (местами более, чем наполовину). Предложен даже специальный термин — ларватон, введенный для обозначения планктона, образованного пелагическими личинками донных видов. Численность личинок одних балянусов может достигать 30000 экз./м3 (Ржепишевский, 1963). В пределе зоопланктон может состоять из личинок бентоса почти на 100%. Например, у Калифорнии в отдельных ячейках апвеллинга зоопланктон может на 90% биомассы состоять из пелагической стадии нектобентической декаподы-галатеиды Pleuroncodesplaniceps, которая даёт до 85% потребления фитопланктона (Longhurst, 1998). Влияние бентоса на пелагиаль не ограничивается поставкой личинок. Большинство пелагических неритических видов в той или иной степени используют дно. Так, существует мнение (Заика, Макарова, 1986), что число медуз Aurelia aurita, являющихся одним из наиболее существенных компонентов экосистемы пелагиали Чёрного моря, прямо зависит от количества зимующих полипов (ведущих прикреплённый образ жизни), способных приступить к стробиляции (отпочковыванию медуз). Происходящие в настоящее время изменения в Чёрном море показывают тесную взаимосвязь бентических и неритических видов.
По размерам в планктоне выделяют: 0,02—0,2 мкм — фемптопланктон, 0,2—2 мкм — пикопланктон, 2—20 мкм — нанопланктон, 20—200 мкм — микропланктон, 0,2—2 мм — мезопланктон, 2—20 мм — макропланктон и >20 мм — мегапланктон. Микропланктон и более крупный планктон (20 мкм—20 мм) называют также сетной планктон, поскольку эти размерные фракции достаточно адекватно облавливают планктонные сети.
Основу фитопланктона создают диатомовые и зелёные водоросли, динофлагелляты и цианобактерии (их часто продолжают называть синезелёными водорослями). Лишь в некоторых районах, например, в Саргассовом море, большую часть продукции создают макрофиты, создающие биомассу до 4—5 г/м2.
Основные группы зоопланктона — Calanoida и эуфаузииды, дающие в сумме в открытом океане более 2/3 биомассы зоопланктона (по органическому углероду), лишь в неритических районах их роль снижается до 1/2 биомассы зоопланктона. Биомасса копепод и эуфаузиид в низких широтах примерно равна, в умеренных и высоких широтах биомасса копепод на порядок больше биомассы эуфаузиид (Longhurst, 2007).
У голопланктонных видов можно выделить следующие части ареала: основа ареала, область размножения и стерильная область выселения.
Поскольку планктон не способен к существенному перемещению по горизонтали, при смешении водных масс всегда происходит смешение и планктона, в них парящего. Поэтому в районах смешения водных масс население имеет смешанный характер.
Важнейшей особенностью пелагиали является полное отсутствие каких-либо неподвижных субстратов. Причём именно неподвижных, поскольку в пелагиали уже более сотни миллионов лет есть многочисленные плотные, но подвижные субстраты — стволы деревьев, а также организмы нектона, на которых формируется своя биота.
Особой разновидностью планктона является нейстон, или плейстон. Настоящие нейстонные виды (обитающие в приповерхностном слое воды или на её поверхности постоянно) есть только в тропических биомах, хотя виды, имеющие нейстонные стадии, есть и в других биомах.
Краткое описание гидрологии Мирового океана дано в Главе 5.
Типы ареалов пелагических видов
У океанических видов основу ареала образуют основные крупномасштабные океанические круговороты. Ареалы многих океанических видов часто охватывают несколько круговоротов. Эти круговороты в значительной степени замкнуты, потери их населения за счёт обмена водой с другими водными массами невелики. Вертикальные миграции океанических видов, по-видимому, существенно не влияют на пространственную структуру популяций, поскольку они не выходят за пределы поверхностных и совпадающих с ними по направлению подповерхностных течений. При этом можно выделить две группы видов: приуроченные к центральным частям круговоротов и к их периферии. В центральных частях круговоротов господствуют олиготрофные условия, трофность возрастает к периферии. Больше ресурсов позволяют существовать большему числу видов.
Для большинства океанических видов фитопланктона характерны ареалы, совпадающие с климатической зоной и охватывающие все океаны в её пределах (Longhurst, 2007) (Рис. 6.1) или охватывает и аналогичный пояс(а) в другом. Типичные ареалы видов планктона показаны на Рис. 6.2.
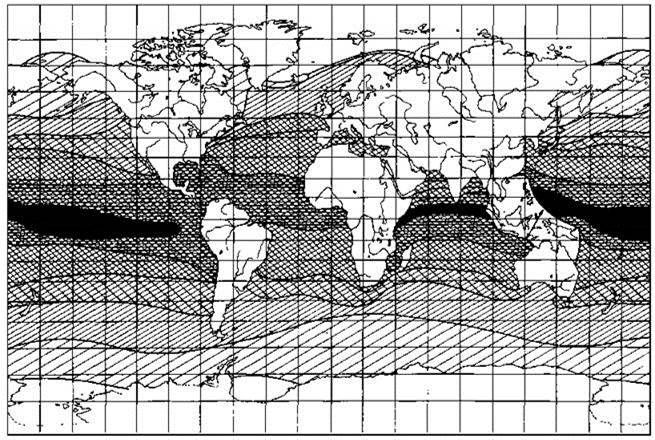
Рис. 6.1. Схема поясного распределения широтных ареалов непрерывного типа. Разные штриховки соответствуют подразделениям в Табл. 6.1. По Ванд-дер-Спуль (1988).
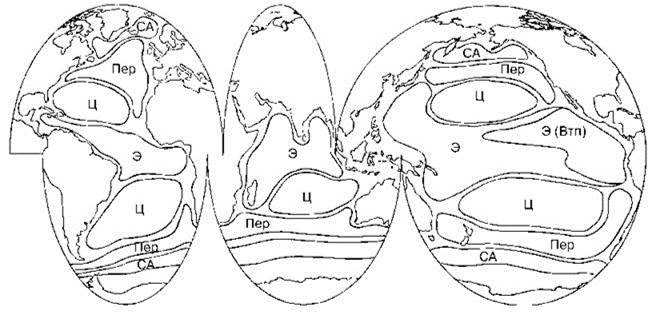
Рис. 6.2. Схема типичных ареалов видов планктона (за исключение полярных). По Longhurst (2001).
Отдельные виды способны встречаться за пределами этих ареалов, иногда в массе, но не способны к созданию независимых популяций. Ареалы: СА — суб(ант)арктический, Пер — переходный, Ц — центральный, Э — экваториальный, Э(Втп) — экваториальный восточнотропический пацифический.
Табл. 6.1. Поясные ареалы. Примеры самых распространенных поясных ареалов и типичные представители. По: Ванд-дер-Спуль (1988).
|
ареал |
широтный пояс |
пример вида |
|
Сплошные в обоих полушариях |
||
|
космополитические |
75°N - 75°S |
Rhyzosolenia alata |
|
ограниченно космополитические |
60°N - 60°S |
Stylocheiron maximum |
|
ограниченно космополитические с тропической субмергенцией |
60°N - 60°S |
Sagitta planotonis |
|
тепловодные |
50°N - 45°S |
Clausocalanus paupulus |
|
тропическо-субтропические |
45°N - 35°S |
Euphausia tenera |
|
тропические |
30°N - 20°S |
Euphausia pseudogibba |
|
экваториальные |
10°N - 10°S |
Clio convexa |
|
Сплошные в одном полушарии |
||
|
полярные |
50°N - 90°N |
Calanus glacialis |
|
субполярные |
50°S - 65°S |
Euphausia triacanta |
|
переходные |
40°S - 50°S |
Euphausia luce ns |
|
(холодно)-умеренные |
30°S - 45°S |
Limacina helicina rangi |
|
Биантитропические |
||
|
бисубполярные |
выше 40°N и S |
Globorotalia pachyderma |
|
двупереходные |
20 - 50°75°N и S |
Thysanoessa gregaroides |
Граница между неритическими и океаническими районами, по-видимому, совпадает с фронтом края шельфа. Эта граница в умеренных широтах наиболее отчётлива во время цветения фитопланктона. Зимой она менее резка и даже может вовсе отсутствовать. Это связано с тем, что большинство неритических видов фитопланктона сохраняется в этот период в виде покоящихся на дне спор, уменьшается также число видов и обилие зоопланктона, отсутствует ларватон. В результате доминирующими становятся не имеющие покоящихся стадий океанические виды, для которых неритические районы — стерильная область выселения.
Неритические виды населяют водные массы шельфа. Многие неритические виды для прохождения полного жизненного цикла нуждаются в дне. У таких видов постоянство ареала обеспечивают донные стадии.
Детальный анализ распределения зоопланктона показал, что среди неритических ЦС следует выделять ЦС прибрежных водных масс и вод открытого шельфа (Шунтов, 2001: 214). Это различие, как и в бентосе следует считать биогеографическим, что означает необходимость разделить неритические виды и ЦС на собственно неритические и прибрежные. В прибрежных ЦС разделение пелагиали и бентали вряд ли целесообразно.
“Внешние условия в стациях неритических и дальне-неритических видов характеризуются, в частности, сравнительно высокой продуктивностью и малой сбалансированностью продукционного цикла” (Беклемишев, 1969: 32).
К.В. Беклемишев (1969) выделил ещё одну группу видов — дальне-неритические виды (Рис. 6.3). Дальне-неритические виды, в отличие от большинства неритических, в своих жизненных циклах не связаны с дном. Структура ареала дальне-неритических видов сходна с ареалами тех неритических видов, которые не связаны с дном, в обоих случаях есть много независимых популяций, поддерживающих довольно многочисленные зависимые популяции. Разница неритических и дальне-неритических в том, что ареалы последних простираются в океан гораздо дальше. С другой стороны, по мнению Ван-дер-Спуля (1988), часть видов, отнесённых Беклемишевым к дальне-неритическим, следует считать океаническими. Они образуют независимые популяции в нейтральных областях глобальных океанических круговоротов. Дополнительно независимые популяции могут существовать в районах со сложной системой течений (например, в Индо-Малайском архипелаге), где могут существовать постоянные круговороты. Возможно, дальне-неритические виды — пограничные.
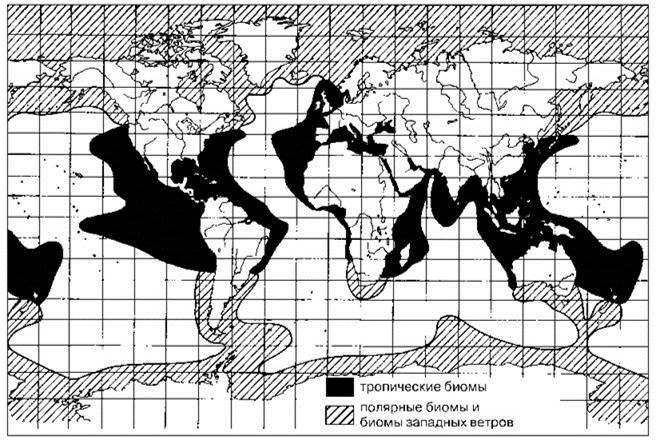
Рис. 6.3. Ареал дальне-неритических видов. По: Ванд-дер-Спуль (1988).
Ареалы ледовых (их также называют ледово-неритические) видов, как и неритических и дальне-неритических включают неритические районы, однако их распространение связано не с дном, а со льдом. Такие виды есть как в Арктике (Рис. 6.4), так и в Антарктике, например род рыб антарктических широколобиков Pagothenia.
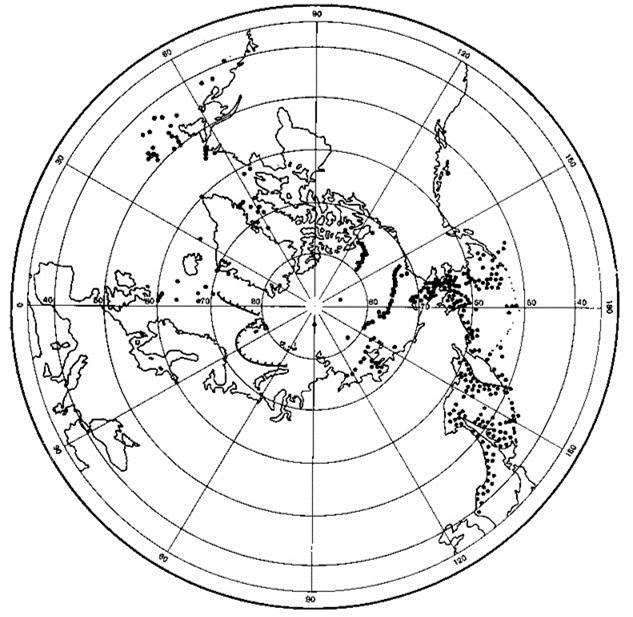
Рис. 6.4. Ареал ледово-неритического вида Calanusglacialis. По Беклемишеву (1969).
Поскольку ареалы меромиктических видов определяет биогеография бентали, они ниже не рассмотрены.
Организмы нектона способны к существенным перемещениям по горизонтали и вертикали и активно выбирают своё место в пространстве. Нектон образуют немногие виды: некоторые кальмары Oegopsida, хрящевые рыбы, немногие костные (большая часть костных принадлежит к планктону), пресмыкающиеся, млекопитающие (киты, ластоногие, каланы, сирены) и многие птицы. Большинство костных рыб и некоторые головоногие имеют планктонные стадии и на ранних стадиях развития временно входят в состав планктона. Рыбы, относящиеся к нектону, достигают крупного размера: хрящевые до 18 м, костистые (меч-рыба, марлин, тунцы) до 3—5 м и до полутонны весом. Они способны плавать со скоростью до 100 км/час. Самые мелкие нектонные виды (сайра Cololabis saira, летучие рыбы Exocoetidae, Oxyporhamphus) имеют длину всего 15—30 см. Представители нектона имеют обтекаемое тело, часто с удлинённым передним концом (типа меч-рыбы).
Многим видам нектона свойственны протяжённые миграции. В этом случае в их ареале часто можно выделить нагульную и нерестовую части ареала. Различие структуры ареалов планктона и нектона иллюстрирует Рис. 6.5. Нагульная и нерестовая части ареала могут совпадать или быть разнесены в пространстве, в предельных случаях нерестовая часть ареала расположена в пресной воде (у осетровых, лососёвых и некоторых других таксонов) или даже на суше (рептилии, ластоногие и птицы). Миграционные пути при этом могут быть за пределами областей как размножения, так и нагульной. У одних видов миграции происходят в меридиональном направлении, часто параллельно берегу; особенно выражены такие миграции у тропических видов, некоторые из которых способны далеко проникать в бореальные (нотальные) регионы в период максимального прогрева их вод. У других видов миграции происходят перпендикулярно берегу. Различают анадромные виды, размножающиеся у берега или даже в пресных водоёмах (лососёвые, осетровые и др.), а нагуливающиеся в морской и катадромные виды, наоборот, размножающиеся в открытых районах, а нагуливающиеся у берегов или даже в пресных водоёмах (наиболее известны лептоцефальные стадии многих видов угрей). Более подробно виды рыб, обитающие в пресной и морской воде рассмотрены в Главе 4. Что касается стерильной области выселения, то, хотя теоретически она и имеется у нектонных видов с планктонным развитием, существенного значения она не имеет и ею можно пренебречь.
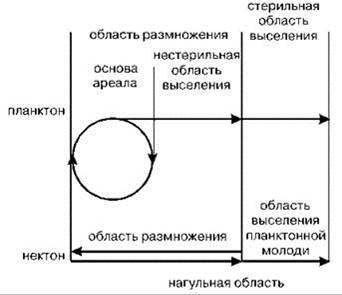
Рис. 6.5. Схема функционального разделения ареалов пелагических животных. По Парину (1968).
Стрелки — направление активных миграций или переноса особей течениями.
Распространение неспособных к полёту птиц (пингвины, истреблённые бескрылые гагарки и бакланы), очевидно, не отличается от распространения прочего нектона. Впрочем, и для способных к полёту морских птиц характерны многие черты, общие с нектоном. Так, среди птиц можно выделить прибрежные, неритические и океанические виды. Прибрежные виды (нырковые утки, гагары, водорезы, большинство бакланов Phalacrocoracidae, некоторые чайки Laridae, крачки Sternidae, пингвины Spheniscidae) постоянно живут (и мигрируют) в узкой (4—5 миль) прибрежной полосе (прибрежной водной массе). Ареалы неритических видов (поморники Stercorariidae, ныряющие буревестники Pelecenoididae, большинство чистиковых Alcidae, морских чаек, крачек, пингвинов, олуш, фрегатов Fregatidae) охватывают весь шельф и сопредельные районы с большими глубинами (неритические и дальне-неритические виды). Океанические виды с прибрежными районами и берегом связаны только в период размножения. К ним принадлежат все альбатросы Diomedeidae и качурки Hydrobatidae и большинство буревестников Procellariidae и фаэтонов Phaethontidae, некоторые крачки. Прибрежные и неритические птицы — преимущественно ныряющие, тогда как океанические добывают пищу исключительно у поверхности.