Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
О сукцессиях - Дополнительные замечания - Био-экология, или синэкология
Существование термина оправданно только в том случае, когда он становится ярлыком для какого-либо явления. Термин “сукцессия” в последнее время используют применительно к любым изменениям: от изменения размерной структуры популяции до изменений, происходящих в геологическом времени, например, при превращении рифтовых озёр в океаны (Старобогатов, 2007). При таком расширенном понимании термина “сукцессия” он становится синонимом слов “изменение” и “смена” для придания тексту наукообразности, т. е. само существование термина теряет какой-либо смысл. Поэтому необходимо кратко рассмотреть разнообразные явления, к которым применяли этот термин, и попытаться понять, какие из них достаточно специфичны, чтобы имело смысл использовать термин. Т. е. в каком случае термин “сукцессия” не может быть заменён более широко распространённым словом без ущерба для смысла. В том числе попытаться разобраться и в значениях различных прилагательных, используемых в сочетании с этим термином.
“Есть ещё один ряд сукцессионных изменений. Озёра, расположенные в рифтовых долинах с течением времени... могут превратиться в океаны” (Старобогатов, 2007: 34).
Очевидно, что при описании изменений размерной структуры популяции или изменений, происходящих в геологическом времени, вполне можно обойтись словом “изменение”. Вряд ли следует называть сукцессией и циклические сезонные процессы. Например, ранней весной в лесах до распускания листвы и в степях появляются многочисленные растения-эфемероиды, быстро заканчивающие жизненный цикл, после чего их надземные части отмирают и остаются лишь подземные покоящиеся стадии (клубни, луковицы и т. п.). Примером таких эфемероидов являются тюльпаны и нарциссы. Говорить о сукцессии здесь нельзя, поскольку из года в год наблюдают одни и те же события, и каких-либо направленных изменений не происходит. В ботанике для таких явлений есть термин сезонный аспект, который и следует использовать в данном случае.
Особый случай представляют смены сообществ, образованных видами с жизненным циклом существенно меньше года. При этом происходят закономерные, повторяющиеся из года в год смены ассоциаций в результате их жизнедеятельности. Такие смены происходит в микробентосе (Бурковский, 2006) и фитопланктоне (Ильяш, Житина, Фёдоров, 2003). В масштабе данных ассоциаций подобные смены можно считать сукцессиями. Но если рассматривать всю ЦС в целом, т. е. сменить масштаб рассмотрения и учитывать не только микробентос, а весь бентос, не только фитопланктон, а пелагическую ЦС в целом, то такие смены следует интерпретировать как смены сезонных аспектов одного и того же не меняющегося, а лишь “колеблющегося” в эволюционно заданных пределах набора ассоциаций. Т. е. целесообразность применения термина “сукцессия” в данном случае зависит от масштаба рассмотрения.
Наиболее часто термин “сукцессия” используют для смен, происходящих в сообществах. Чтобы разобраться в многообразии возможных смен предложено много классификаций на разных логических основаниях (см., например, Миркин и др., 2002): по масштабу времени (быстрые, средние, медленные, очень медленные), по обратимости (обратимые и необратимые), по степени постоянства процесса (постоянные и непостоянные), по происхождению (первичные и вторичные), по тенденциям изменения продуктивности (прогрессивные и регрессивные), по тенденции изменения видового богатства (прогрессивные и регрессивные), по антропогенности (антропогенные и природные), по характеру происходящих во время сукцессии изменений (автотрофные и гетеротрофные). По мнению А.П. Шенникова (1964: 319), подобные классификации — это “классификации смен по их причинам”. В действительности, как указывает С.М. Разумовский (1999: 345), это классификации причин, но не смен, поскольку одно дело причина, вызвавшая процесс, а совсем другое — сам процесс. Связь тут далека от однозначной функциональной. Нередко такие классификации делают на основе произвольно выбранных показателей, которые могут меняться во время сукцессии.
Рассмотрим в качестве примера подразделение сукцессий на первичные и вторичные, введённое F. Clements (1916). Под первичными понимают сукцессии, начинающиеся с момента образования биотопа (например, после выкапывания пруда), под вторичными — сукцессии в местах, где уже было сообщество (например, после заливания ранее осушённого пруда или на вырубках). Первичные сукцессии — это всегда сукцессии экогенетические. Что же касается вторичных сукцессий, то тут возможны два варианта:
— если в результате воздействия были разрушены и сообщество, и его биотоп, то вторичная сукцессия — это экогенетическая сукцессия, начинающаяся с той точки, которой соответствует степень разрушения биотопа (лес может быть только вырублен или на его месте выкопан пруд), т. е. отличаться от первичной только точкой начала, положение которой определяет степень внешнего воздействия.
— если внешнее воздействие привело к разрушению только сообщества, оставив нетронутыми сделанные ими преобразования биотопа (лес вырублен без нарушения почвенного покрова), то в таком случае вторичная сукцессия — это демутационная сукцессия.
Очевидно, что вторичные сукцессии в представленных двух вариантах будут разными, и тем более чем более развиты демутационные сукцессионные ряды в ареале данной ЦС. С другой стороны, в первом варианте первичные и вторичные сукцессии — синонимы. Иными словами, процессы, отнесённые по данной классификации к различным категориям (первичные или вторичные сукцессии), в действительности могут быть как разными, так и идентичными, что говорит о том, что деление сукцессий на первичные и вторичные не выявляет их биологической специфики, является биологически неинформативным. Это, впрочем, и неудивительно, поскольку логическим основанием для деления здесь является не свойства изучаемого процесса, а интенсивность внешнего воздействия, причём такая интенсивность оценивается не по реакции изучаемого объекта на воздействие, а из априорно заданных представлений.
Другой пример: подразделение сукцессий на автотрофные и гетеротрофные. Подобное подразделение имеет в своём основании идею о том, что отношение первичной продукции (Р) к дыханию (R) в ходе сукцессии “приближается к единице (равновесию)” (Одум, 1986: 167). Автотрофные сукцессии — это сукцессии, начинающиеся с сообществ, у которых P/R>1, гетеротрофные — P/R<1, при том в гетеротрофной сукцессии “стационарного состояния достичь невозможно” (Одум, 1986: 167).
Представление о том, что P/R=1 — это состояние равновесия, является экстраполяцией представлений экологов, изучавших некоторые наземные экосистемы, на всю биосферу. Каждое сообщество при этом отождествляют с закрытой системой, основным, если не единственным, источником энергии в котором является фотосинтез входящих в состав сообщества растений. Между тем любое сообщество — открытое, что особенно относится к водным сообществам, где доля аллохтонного органического вещества, энергию которого использует сообщество, достигает 100%. У водных сообществ за пределами фитали (за исключением сообществ хемобиоса) вообще нет первичной продукции. Нередко и в пределах фитали первичная продукция очень незначительна, например, во многих сообществах рыхлых грунтов. Однако это не мешает им находиться в состоянии равновесия миллионы лет. Поэтому соотношение первичной продукции и дыхания как показатель устойчивости применительно к ним оказывается совершенно неинформативным. Теория, применимая лишь к ограниченному числу объектов, не может претендовать на роль общей. Чтобы не менять теорию, такие сообщества часто объявляют неполноценными. С моей точки зрения, странно считать сообщества, населяющие больше половины поверхности Земли, неполноценными. Логичнее неполноценной назвать теорию. Если же придерживаться представлений, описанных Одумом, то к первичным продуцентам следует отнести любой низший трофический уровень, поставляющий энергию в ЦС, в том числе и потребителей детрита, хотя это и кажется непривычным. Более того, если начать копать глубже, то принятая в настоящее время практика не вполне последовательна. Сейчас к первичным продуцентам относят “автотрофные организмы, в основном зелёные растения, которые могут производить пищу из простых неорганических веществ” (Одум, 1986: 29), т. е. получающие энергию для жизнедеятельности и синтеза нового органического вещества за счёт любых источников, кроме окисления другого органического вещества. При таком определении вестиментиферы чёрных курильщиков, использующие энергию окисления восстановленных соединений металлов, являются автотрофами (хемоавтотрофами). Тот же вид, живущий на трупе кита или затопленном сухогрузе, использующий энергию окисления органического вещества, становится уже гетеротрофом. В любом случае отношение Р/R и деление сукцессий на автотрофные и гетеротрофные имеют ограниченное применение. Классический пример гетеротрофной сукцессии — динамика простейших в сенном настое (Woodroff, 1912) — в действительности является нормальным процессом потребления органики редуцентами, который можно отождествлять с сукцессиями в природных сообществах разве что в программе средней школы.
Некоторые из предложенных классификаций основаны на изменении свойств сообществ (продуктивности, видового богатства и др.), действительно наблюдавшихся в ходе отдельных сукцессий. Эти изменения безосновательно сочли универсальными, несмотря на то, что в ходе других сукцессий те же свойства менялись иначе, порой диаметрально противоположным образом.
Поэтому множественность классификаций сукцессий, проводимых практически по любому из мыслимых логических оснований, оправдана только с точки зрения континуалистов, считающих сукцессии стохастическим процессом конкурентного взаимодействия отдельных случайно обитающих вместе видов, происходящим под действием разнообразных внешних факторов. Очевидно, что, в зависимости от целей исследователя, подобные классификации можно строить на любом основании, а число их можно увеличивать до бесконечности. Так, П.Д. Ярошенко (1950: 57) вполне логично указывал на необходимость разделения антропогенных смен на:
— смены в социалистических странах;
— смены в капиталистических странах.
В то же время, очевидно, что подобные классификации мало дают для понимания эндогенной динамики сообществ.
С точки зрения изучения динамики естественных сообществ, имеет смысл разделить смены на экзогенные и эндогенные. Движущей силой эндогенных смен является функционирование ассоциаций. Движущей силой экзогенных смен может быть что угодно, кроме функционирования ассоциаций. Термин “сукцессия” целесообразно применять только к эндогенным сменам, экзогенные же смены следует называть просто “сменами”. Необходимо подчеркнуть, что большая часть внешних воздействий своим результатом имеет ситуацию сходную с той, что встречается или уже встречалась в пределах ареала ЦС. Т. е. экзогенные воздействия искусственно приводят к смене существовавшего в данном месте сообщества другим сообществом этой же ЦС. После прекращения внешнего воздействия начинается эндогенная сукцессия, ничем не отличающаяся от обычной. Так, рубка леса с точки зрения ЦС (судя по её реакции) эквивалентна верховому пожару или ветровалу; хищническая рубка, проводимая с применением тяжёлой техники, с разрушением почвенного покрова — низовому пожару. Экзогенное воздействие может даже совпадать с сукцессией. Так, в сукцессии березняк → ельник выборочная рубка берёзы только ускорит прохождение сукцессии.
“Антропогенные смены растительного покрова в условиях нашего социалистического общества несут принципиально отличный характер от смен растительного покрова в условиях капитализма” (Ярошенко, 1953: 126).
Следует также разделять сукцессии в ЦС и смены в группировках видов, в том числе — антропогенных. В ЦС сукцессии — исторически сложившийся жёстко детерминированный процесс. Смены же в группировках — стохастический процесс преобразования одной случайной группировки в другую. Одна из основных ошибок континуалистов состоит в смешивании этих двух принципиально разных процессов и перенесение закономерностей, выявленных на случайных группировках (которые легко воспроизвести в лабораторных условиях), на ЦС (Рис. 1.55). Впрочем, некоторые, а иногда и многие процессы сукцессий в ЦС и группировках могут совпадать. В то же время между ними имеются и большие принципиальные различия. Возможно, правильнее было бы признать существование двух разделов экологии: экологии природных естественных ЦС и экологии антропогенно модифицированных, созданных или поддерживаемых человеком группировок видов. Необходимо, конечно, помнить, что идеальная ЦС — это предел, к которому стремится биота. Реальные ЦС обладают в той или иной степени чертами группировок и могут возникать в результате эволюционного преобразования группировок и входящих в них ценофобных видов.
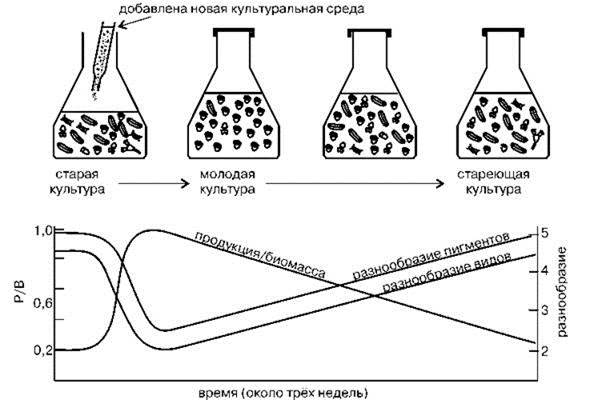
Рис. 1.55. Модель сукцессии. По Одуму (1968) из Нинбурга (2005).