Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Метод соответствия ранга биогеографического региона рангу эндемов - Методы группирования регионов - Методы районирования - Общая биогеография
Естественная классификация должна исходить из свойств классифицируемых объектов. Географическая и биологическая биогеографии изучают свойства разных объектов, поэтому результаты географического и биологического биогеографического районирования различны. Хотя методы географической биогеографии в этом разделе (как и в прочих) не рассмотрены, широкое распространение в биологической литературе результатов, полученных с их помощью, вынуждает сказать несколько слов об их биологическом содержании.
Изучение конкретного распределения сообществ по поверхности Земли выявляет сходные, а порой и совершенно идентичные сообщества в весьма удалённых районах. В географической биогеографии используют это физиономическое сходство территорий и сообществ. Оценка сходства — первая причина возникновения субъективности. Кроме того, как следует из теории структурализма, отдельные ассоциации, из которых состоят сообщества, есть лишь пространственно-временная форма существования ЦС. Ассоциации связаны динамически, плавно или не очень переходят одна в другую в ареале ЦС. Наличие стадий, переходных между ассоциациями, а также мелкая мозаичность, ведущая к тому, что в одно сообщество часто включают разные ассоциации, заставляет выделять при физиономическом подходе весьма широкие переходные зоны. Сообщества, которые и после этого не удаётся уложить в прокрустово ложе заготовленной схемы, относят к интра- и экстразональным.
“Окончательное разделение лесного пояса Европейской России и сопредельных территорий на зоны лесостепи и широколиственных лесов, южной, средней и северной тайги происходило в течение позднего голоцена, в основном в 1—2 последних тысячелетий” (Турубанова, 2002: 15). “Судя по археологической и исторической литературе, <это разделение> в значительной степени было обусловлено всё усиливающейся хозяйственной деятельностью” (Турубанова, 2000).
Например, в географической биогеографии по физиономическому сходству сообществ выделяют зону хвойных лесов (= тайга, бореальные леса) и зону широколиственных лесов (= неморальные леса), между которыми выделяют зону или подзону смешанных лесов (см., например, Рис. 2.26). Структуралисты же выделяют зону широколиственных лесов, где эдификаторы климакса — разные виды дуба Quercus и зону тайги, где эдификаторы климакса — глосеменные (ели, пихты или тёмнохвойные сосны) и не выделяют зону смешанных лесов. Они объясняют существование смешанных лесов тем, что в ходе сукцессий существуют стадии, когда имеется только травянистая растительность, стадии с доминированием лиственных и хвойных деревьев (Рис. 1.38, Рис. 2.25). Поскольку смена стадий сукцессий происходит не мгновенно, имеются и переходы между ними. В одних случаях лиственный древостой ещё не выпал, а хвойный уже подрос, в других — хвойный древостой ещё не выпал, а лиственный уже подрос. Из этих двух схем видно, что хвойные деревья могут сменять и мелколиственные, и широколиственные, мелколиственные могут сменять широколиственные и наоборот. Поэтому, с точки зрения структуралистов, “смешанные” леса являют собой хороший пример экотонов ЦС с широколиственными или хвойными доминантами климакса, где переход происходит между ассоциациями с хвойными и лиственными доминантами древесного яруса. Факт преобладания в отдельных районах этих ЦС лиственных лесов или заметная примесь хвойных в лиственных лесах является не особенностью данных ЦС, а результатом частых нарушений, большей частью антропогенных (рубок, пожаров и т. п.). В частности, климаксной ассоциацией в нечернозёмной России на север до Карелии (Рис. 1.36, Рис. 2.27) является ассоциация с доминированием дуба черёшчатого (Quercus robur). И именно дуб следовало бы считать характерным для Руси, да так раньше и было. Например, А.С. Пушкин в “Руслане и Людмиле” описывает путешествие главного героя по Европейской России от юга до севера. При этом он упоминает дуб 14 раз, а березу всего 3, причём один из них в словосочетании “березовая дубрава”. Однако к настоящему времени дуб почти повсеместно вырублен. Особенного размаха вырубка достигла в конце XIX в., и места, ранее занятые дубравами, теперь заняты ельниками, сосняками, березняками и т. п., но особенно полями, поскольку почвы и режим увлажнения на территориях, ранее занятых климаксом, наиболее благоприятны для пашен (Советов, 1867). На северной границе ареала дуба, несмотря на возобновление дуба в ельниках (местами интенсивное), дубравы отсутствуют. Отдельные дубы остались лишь в местах, откуда неудобно вывозить срубленные деревья (долины рек, особенно крутые склоны) (личн. наблюдения И.Ю. Попова). Эта деятельность человека и создаёт обманчивое впечатление интразональности, широко обсуждаемое в литературе. Аналогичные изменения произошли и в Северной Америке. Антропогенный характер имеет и южная граница ареалов хвойных деревьев. Фактически ещё 2500—500 лет назад ареалы широколиственных (дуб, бук, клён, ясень, липа, вяз, граб, лещина) и тёмнохвойных (ель Picea abies и пихта Abies spp.) деревья в Восточной Европе практически совпадали (подробнее см. Главу 3 и Рис. 3.63), т. е. к зоне смешанных лесов в Восточной Европе следовало бы отнести всю территорию, при этом не было бы ни зоны широколиственных лесов, ни тайги.
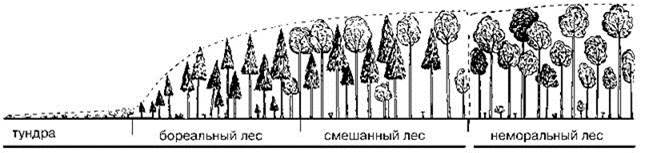
Рис. 2.26. Часть градиента экоклины тундра — тропики. По Уиттекеру (1980).
“На обширных пространствах Русской равнины самые северные местонахождения дуба — в долинах рек; по ним он нередко проникает на 200—300 км севернее, чем по междуречьям. Г.Ф. Морозов (1914) объяснял это лучшим прогреванием пойменных почв и их меньшей оподзоленностью; по мнению А.К. Денисова (1980) причины — в более быстром размерзании и более высокой температуре зимой пойменных почв, а также в большей влажности и близости грунтовых вод” (Восточно-Европейские леса..., 2004: 119—120).
Поэтому, если уж выделять зоны смешанных и мелколиственных лесов, то правильнее их назвать, например, “область широколиственных лесов и тайги с интенсивными антропогенными нарушениями” и не пытаться искать её биологического смысла. Антропогенные нарушения, хотя и сильно варьируют во времени и пространстве, происходят сравнительно плавно (даже в настоящее время скорость их протекания сопоставима с продолжительностью жизни поколения Homo sapiens) и охватывают огромные площади, создавая обманчивое впечатление естественности (см. также Главу 3).
Справедливости ради надо отметить, что зону смешанных лесов выделяют отнюдь не все биогеографы. Нет её, например, у И.И. Пузанова (1938).
“Миф белой берёзы — это искусственный, поэтический миф, причём сравнительно недавний... До конца девятнадцатого века о берёзах никто и не думал, их даже официально не считали белыми. У Рылеева стволы берёз мрачно чернеют, а у Мережковского даже “чернеют как уголь”. Поклонение Белой Берёзе идёт главным образом от Есенина” (Метелица, 2004: 8). Такая смена представлений о берёзе — следствие возрастания интенсивности рубок и других антропогенных воздействий, а также сокращения площади пашен и других сельхозугодий (Сушков, 1974). И то, и другое привело в конце XIX в. к резкому увеличению лесов на ранних стадиях сукцессий, т. е. возросло и число берёз (эдификатора ранних стадий сукцессий после рубок и пожаров) вообще, и молодых берёз с белой корой в особенности.
“Боры из обыкновенной сосны Pinus sylvestris с подлеском из брусники Vaccinium vitis-idaea и напочвенным покровом из Шреберова плеврозия Pleurozium schreberi встречаются от Кольского полуострова до Среднего Дона. По облику и видовому составу это сообщество из группы формаций бореальных хвойных лесов. Поэтому боры-брусничники Среднего Дона следовало бы трактовать как экстразональную растительность, как далёкий южный эксклав светлохвойной тайги, существование которого вблизи южной границы летнезелёных лесов умеренной зоны должно вызывать глубокое изумление. Однако на Кольском полуострове, в подзоне северной тайги, бор-брусничник представляет собой заключительную стадию ксеросерии, которая сменяет стадию вересковой пустоши из обыкновенного вереска Calluna vulgaris и преобразуется в ходе последующей сукцессии в климаксный ельник-брусничник с 2 ярусами: древесным ярусом из европейской ели Picea abies и кустарничковый ярус из брусники Vaccinium vitis-idaea. На Среднем Дону такой же бор оказывается тоже стадией ксеросерии, но не последней, а предпоследней. Он возникает на месте бора из обыкновенной сосны Pinus sylvestris без подлеска, но с травяным ярусом из поникшего перловника Melica nutans и, в свою очередь, сменяется бором, имеющим также только древесный и травяной ярусы. Однако последний сложен майским ландышем Convallaria majalis” (Тимонин, Озерова, 2002: 110) (Рис. 2.27).
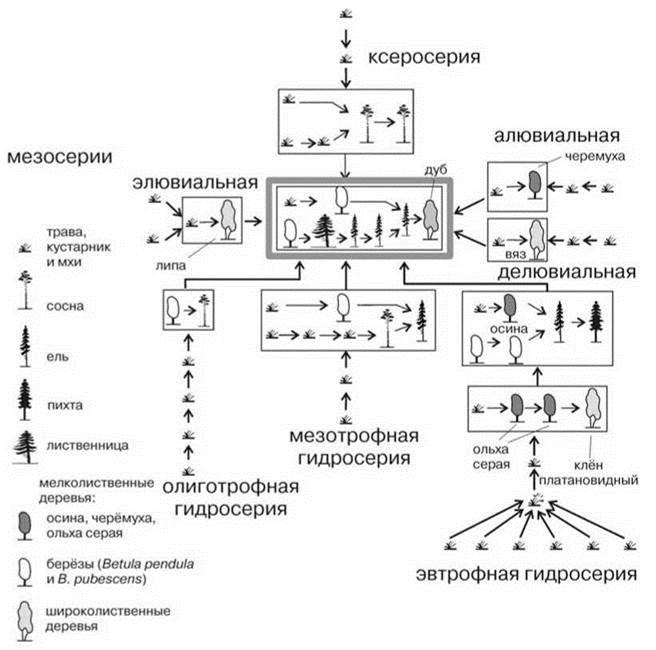
Рис. 2.27. Сукцессионная система Ветлужского ботанико-географического района. По С.М. Разумовскому (1981).
Район расположен северо-восточнее Московского и граничит с ним, схема Московского района дана на Рис. 1.38 (их расположение показано на карте на Рис. 3.65). Расшифровку терминов и обозначений см. на Рис. 1.38.
Другой пример: В.Б. Сочава, Т.И. Исаченко и А.Н. Лукичева (1953) выделяют на территории Западной Сибири “западный меридиональный сектор”, характеризующийся преобладанием сосняков. Авторы объясняют это климатом (близость Уральского хребта снижает количество осадков) и преобладанием песчаных почв. На самом деле в этом районе песчаные ледниковые (в данном случае — единственно возможная песчаная материнская порода) отложения практически отсутствуют, а резкое преобладание сосняков, действительно почти сплошь покрывающих водоразделы, связано с постоянным выжиганием лесов местными оленеводами (для увеличения площади ягельников) (Разумовский, 2011). Очевидно, что естественный с точки зрения географов выдел, с точки зрения биолога представляет собой лишь временную стадию развития тайги и вовсе не заслуживает какого-либо выделения на биогеографической карте (но вполне может быть показана на карте хозяйственной деятельности человека, наряду с полями и сенокосами).
“Хотя понятия “саванна” и “степь” были введены для обозначения определённых типов растительности, в последнее время они применяются географами скорее для обозначения соответствующих климатических зон... В этом случае, по мнению автора, следовало бы пользоваться чисто климатической терминологией... ботанические термины... должны применяться для характеристики какой-то определённой категории растительного покрова” (Вальтер, 1968: 276).
В биологической литературе из-за неразработанности биологических методов классификации и выделения ЦС и сообществ есть тенденция использовать географические подходы. В начале главы уже обсуждена принципиальная неприемлемость такого подхода для целей биологии. Но такой подход неприемлем и по другим причинам. Желание биологов использовать географические методы и их результаты основано на ошибочном представлении о том, что в географии всё давно разработано, устоялось и можно взять готовую схему и использовать её в биологических целях.
Помимо принципиальной непригодности таких схем, их применение не решает проблему, поскольку общепринятых географических схем нет. Критерии выделения регионов и в географии весьма субъективны. Это следует хотя бы из того, что один и тот же биом разные авторы понимают весьма различно, хотя и называют одинаково. В качестве примера на Рис. 2.28 дано распространение степей по разным авторам. Из сравнения данных схем становится очевидной высокая степень их субъективности, неоднозначности применения этого подхода и в самой географии.
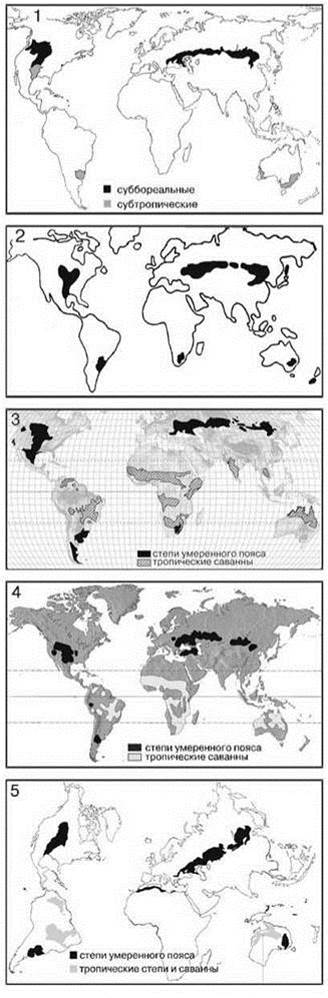
Рис. 2.28. Распространение биома степей по разным авторам.
1 — по: А.Г. Исаченко, А.А. Шляпникову (1989); 2 — по: В.Г. Мордковичу (1982); 3 — Woodward (2008); 4 — Allaby (2006); 5 — по классификации ЮНЕСКО (План действий..., 1984).
Необходимо отметить, что участок биома степей в Южной Америке (пампы), показанный на всех схемах, возможно, не является естественным, а имеет антрогенное происхождение (см. Главу 3).
Та же картина наблюдается и в любом более мелком масштабе, в том числе и в масштабе ландшафтов. Например, болото — вполне ясный и хорошо очерченный, казалось бы, тип ландшафта. Однако даже по официальным данным разных ведомств площадь болот в России различается на 20%: в начале 1998 г. в лесном фонде было 128,3 млн. га (Лесной фонд России, 1999), но по данным государственного учёта земель болота в лесном фонде России занимают всего 101,2 млн. га (Думнов и др., 2001). Неофициальные данные дают ещё больший разброс: до 273,0 млн. га (Ефремов и др., 1994) и даже 361,1 млн. га (Вомперский и др., 1999). Достойно удивления, что при общем разбросе размеров площадей в три раза, все источники считают возможным приводить свои цифры с точностью до четырёх значащих цифр.
Из сказанного выше вовсе не следует, что выделение зон в биогеографии не нужно и бессмысленно. Однако для получения осмысленных с биологической точки зрения зон следует использовать биологические критерии, тем более, что имеются образцы такого районирования с получением биологически значимой информации.
В качестве примера можно привести выделение зон наземной растительности С.М. Разумовским (1999). Начав с установления ботанико-географических провинций, он выявил коренные ассоциации их климаксов, затем сравнил облик древесных эдификаторов климаксных ассоциаций, общее число которых (для всей Земли) составляет лишь несколько десятков, и обнаружил, что “эти виды очень легко и естественно распадаются на четыре различных между собой и очень монолитных морфологических типа:
1. Вечнозеленые перистолистные деревья с почками, лишёнными почечных чешуй.
2. Вечнозеленые широколистные деревья с почками, защищёнными почечными чешуями.
3. Летнезелёные широколистные деревья с почечными чешуями.
4. Вечнозелёные деревья с игольчатыми листьями ксероморфной структуры и с почечными чешуями” (Разумовский, 1999: 73).
Эти зоны и являются естественными биологическими зонами растительности Земли: тропическими, субтропическими и широколиственными лесами и тайгой.
ЦС, входящие в одну зону, могут приобрести физиономическое сходство конвергентно и вообще не иметь ничего общего ни в своём составе, ни в происхождении. Сходная ситуация имеется и в таксономии: в воде активно плавающие животные приобретают форму рыбы, но гораздо информативнее классификация не на основе жизненных форм, а филогенетическая, учитывающая то, что одни из этих “рыб” — рыбы, другие — рептилии, третьи — млекопитающие, четвёртые — птицы, пятые — головоногие моллюски и т. д. Из того факта, что в воде они приобрели сходную форму, мало что можно сказать об их внутреннем строении, биохимии, происхождении и т. п. Но все эти данные вполне доступны из филогенетической классификации, построенной на основе их генетического родства. Аналогично, объединение биогеографических регионов по их происхождению гораздо более информативно для понимания их биологических свойств, тогда как зональные классификации лишь показывают многообразие ЦС, образовавшихся на разной биотической основе, но в сходных абиотических условиях.
Для других целей (например, для оптимального размещения санаториев) более информативными являются иные логические основания для объединения регионов, причём логические основания для разных задач могут быть разными. В некоторых случаях более информативными будут и чисто физиономические основания.
Итоговая схема биогеографического распространения обычно иерархична. Критерии группирования районов могут быть весьма различны. В биологической биогеографии методы группирования регионов можно подразделить на три группы, в зависимости от того, что, по мнению авторов схем они должны отражать. Наиболее распространена точка зрения, согласно которой схемы должны отражать степень эндемизма, или, что то же самое степень сходства и отличия биот. Другие полагают, что схема должна давать представление о древности, времени происхождения, истории расселения и о центрах развития биот. И, наконец, третьи полагают, что схемы должны отражать распределение экосистем. Примеры схем, отражающих разные точки зрения приведены на Рис. 2.29—2.31 и на цветной вклейке Рис. Ц.4. На Рис. 2.29 приведена классическая схема Wallace и Sclater, основанная на оценке фаунистического сходства. Почти сразу же после того, как она была предложена, её подвергли критике за то, что она не даёт представления ни о древности, ни о центрах развития и истории расселения. Для отражения этих аспектов были предложены схемы на Рис. 2.30 и 2.31. И, наконец, распределение ЦС показано на Рис. Ц4. Стремление отразить историю развития биот привело к объединению мало сходных регионов только из-за ареалов древних таксонов, имеющих малое значение в экосистемах регионов, как например, в Архигее (Рис. 2.31). Схема, построенная на основе сходства (Рис. 2.29) ближе к схеме, отражающей распределение ЦС (Рис. Ц.4). Основное их различие обусловлено тем, что в первая построена методом сравнения списков биот, а вторая — методом выделения ЦС.
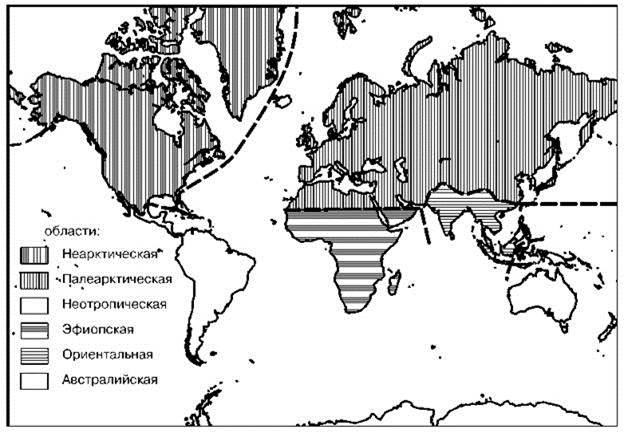
Рис. 2.29. Схема фаунистического районирования суши Wallace и Sclater (по: Huggett, 2004), основанная на оценке сходства биот.
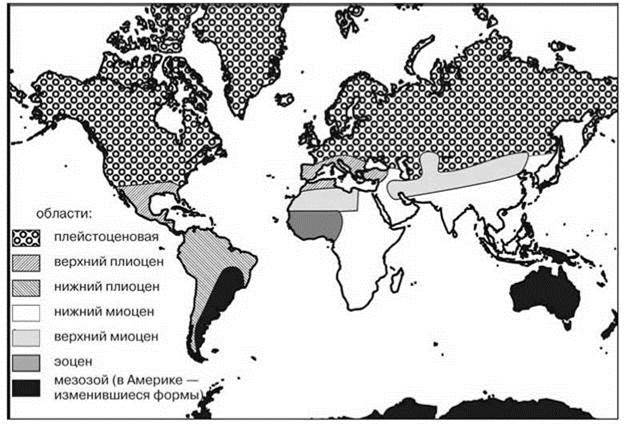
Рис. 2.30. Схема биогеографического районирования суши Rutimeyer (1867), отражающая древность фаун (по Лаптеву, 1940).
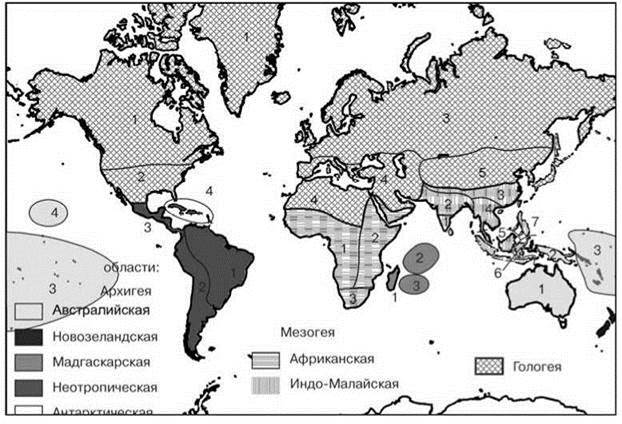
Рис. 2.31. Схема биогеографического районирования суши Бианки (1918), отражающая наличие таксонов, принадлежащих той ли иной эпохе (по Лаптеву, 1940).
Области (подобласти). Жирным шрифтом выделены подразделения ранга более высокого, чем область
Архигея — вторичная суша: Неотропическая (1 — Бразильская, 2 — Патагонская, 3 — Центральноамериканская, 4 — Вестиндская); Мадагаскарская (1 — Мадагаскарская, 2 — Сейшельская, 3 — Маскаренская); Австралийская (1 — Австралийская, 2 — Новогвинейская, 3 — Полинезийская, 4 — Гавайская); Новозеландская; Антарктическая.
Мезогея — третичная суша: Африканская (1 — Западноафриканская, 2 — Восточноафриканская, 3 — Южноафриканская); Индо-Малайская (1 — Южноидийская, 2 — Переднеиндийская, 3 — Китайско-Гималайская, 4 — Индо-Китайская, 5 — Малакско-Зондская, 6 — Целебесская, 7 — Филипинская).
Гологея — четвертичная суша: (1 — Бореоамериканская, 2 — Сонорская, 3 — Боревразийская, 4 — Средиземноморская, 6 — Нагорноазиатская).
Названия высших подразделений основаны на терминологии итальянского геолога Джованни Ардуино, который, изучая геологию северной Италии в 1759 г., разбил шкалу времени на первичный, вторичный и третичный периоды. Позднее к ним добавили четвертичный период. Названия основаны на процентном содержании в отложениях данного времени ископаемых моллюсков, близких к современным видам. Вторичный период соответствует периоду до кайнозоя (>65 млн. лет назад), четвертичный — ледниковому периоду (<1,8 млн. лет назад), третичный — периоду между ними. Т. е. по мнению автора схемы древнейшие таксоны архигеи (однопроходные, сумчатые и неполнозубые) образовались до кайонозоя, мезогеи (низшие плацентарные) — в кайонозое до ледникового периода, гологеи (высшие плацентарные) — во время ледникового периода.
“Все опубликованные до сих пор (т. е. за последние два века) схемы территориального расчленения растительного покрова при всех их различиях объединяются одним общим признаком — все они строились дедуктивно, т. е. “сверху вниз”, от крупных единиц к более мелким. Этот, формально логичный, но биологически ненаучный способ состоит в том, что изучаемое множество (в данном случае совокупность растительных сообществ планеты) произвольно делится на логические классы по субъективно выбранным признакам. При этом степень дробности деления также вполне субъективна и в принципе почти неограниченна, а границы получаемых единиц в сущности не являются вообще границами, т. е. не совпадают с реально существующими в природе изолиниями качественных скачков” (Разумовский, 1980: 11).
В основу группировки регионов следует положить анализ ценофильных видов. Разделение видов на ценофильные и ценофобные требует проведения в каждом биогеографическом регионе серьёзных исследований. В данный момент эти исследования только начинаются и на практике в морской, да и пресноводной и даже наземной биогеографии приходится считать все виды ценофильными, что, без сомнений, неверно. Как уже говорилось, доля ценофобных видов в разных таксонах наземных растений варьирует от 0 до 100%. Это говорит о том, что одни таксоны более пригодны для целей биогеографического районирования, нежели другие, а некоторые — и вовсе неинформативны. Использование ценофобных видов при сравнении биогеографических регионов приводит в первую очередь к сглаживанию различий. Также теоретически возможно, но представляется маловероятным для антропогенно слабонарушенных регионов, что учёт ценофобных видов существенно исказит картину: заставит объединить районы, различающиеся ценофильной биотой и наоборот. В сильнонарушенных регионах, к которым относится большая часть суши, ценофобные виды составляют существенную долю биоты, часто гораздо больше половины. Многие из этих ценофобных видов переселены умышленно или случайно человеком из других биогеографических регионов. Сам факт такого переселения не всегда очевиден, иногда его устанавливают только после тщательных исследований. В то же время многие ценофильные виды, в том числе и игравшие в природных ЦС существенную роль, во многих районах истреблены или их численность существенно снизилась, в результате чего естественный облик ЦС был изменен, порой до неузнаваемости (многочисленные примеры этого приведены ниже, особенно в Главе 3). Можно с уверенностью сказать, что использование при сравнении полных списков биот в таких случаях даст картину очень сильно отличающуюся от той, которая будет получена при сравнении списков конкретных биот, т. е. тех, которые были до прихода человека и которую только и следует считать естественной.
На практике, при проведении районирования необходимо соблюдение правил логики. В первую очередь это значит, что классификацию следует делать на одном логическом основании. Это означает также необходимость соответствия идеологий, положенных в основу методов, использованных для выделения районов и их группировке.
В ботанике существует также разделение биогеографических схем на флористические, геоботанические и ботанико-географические (Камелин, 1990, 2012). Здесь, вслед за Лавренко (1942, 1962) и Разумовским (1999, 2011) мы считаем их синонимами, поскольку ареалы ценофильных видов определяют ЦС, в состав которых они входят, различия флоры отражают то, что разные территории заняты различными ЦС.
Метод соответствия ранга биогеографического региона рангу эндемов
Многие авторы (Тахтаджян, 1970; Старобогатов, 1970; Гурьянова, 1972; Василенко, 1974; Несис, 1982а; Хржановский и др., 1986; Кафанов, Кудряшов, 2000; Абдурахманов и др., 2003, Spalding et al., 2007 и многие другие) предлагают присваивать ранг биотическому региону в зависимости от ранга его эндемов или, по крайней мере, учитывать ранг эндемов при определении ранга биогеографического региона. Посмотрим, как на практике применяют это требование. Проводя биогеографический анализ шельфа Северного полушария, Е.Ф. Гурьянова пишет: “Самая высокая категория — царство; важнейшими критериями для её выделения служит очень высокий ранг эндемизма — наличие эндемичных классов и подклассов, хотя бы и немногих, и большого числа эндемичных отрядов” (Гурьянова, 1972: 11). Далее она сама выделяет два царства. Первое — тропическое — в полной мере удовлетворяет предложенным ею критериям для царства. Второе объединяет обе бореальные и арктическую области. Это царство ни в коей мере не удовлетворяет её же собственным критериям. Аналогичные выводы сделал С.М. Разумовский (1999), разбирая самую популярную в отечественной биогеографической литературе схему фитогеографического районирования Земли А.Л. Тахтаджяна (1970) (Рис. 3.28). Очевидная невозможность применить данный метод на практике даже его энтузиастами однозначно свидетельствует о его несостоятельности.
С другой стороны, показано, что число родов является простой математической функцией числа видов: чем больше видов, тем больше родов (Кафанов, 1991). Иными словами, более богатая видами биота будет содержать и большее число высших таксонов (родов, семейств, отрядов и т. д.), что мы, собственно говоря, и имеем: в тропической биоте больше таксонов высокого ранга, эта разница и есть эндемичные таксоны. Искать в этом какой-либо биологический смысл вряд ли стоит, скорее, такая закономерность “описывает, по-видимому, не реальные биологические закономерности, а эффекты, порождаемые конечной “пропускной способностью” человеческого мозга и неосознанным стремлением исследователей компенсировать это ограничение” (Кафанов, 1991: 41—42). Как сказано выше, Ранг таксона объясняют не генетические, а экологические причины, он функция не времени, а различий в ресурсах, потребляемым исходным и образовавшимися таксонами.
“Голарктическое царство” характеризуется [А.Л. Тахтаджяном] 39 эндемичными семействами. В Московской области из этого числа имеются лишь три семейства: Adoxaceae, Butomaceae и Scheuchzeriaceae**. При этом Butomaceae отсутствует в Северной Америке... Таким образом, даже такая естественная, реально существующая и хорошо известная флористическая единица, как Голарктис, не может быть ни выделена, ни охарактеризована по предлагаемому А.Л. Тахтаджяном принципу” (Разумовский, 1999: 30).