Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Двустворчатые моллюски Арала - Причины и скорость образования новых видов и экосистем - Био-экология, или синэкология
На Аральском море человек в конце прошлого века поставил крупномасштабный эксперимент по образованию новой экосистемы. С резким сокращением, а временами и полным прекращением пресного стока в море, произошло стремительное нарастание солёности и общее уменьшение площади водоёма (Рис. 1.26). До зарегулирования стока рек основными первичными продуцентами являлись макрофиты (Карпевич, 1975), но к 1990-м гг. большинство видов макрофитов вымерло: из 37 сохранилось только 4 (Доброхотова, 1971; Orlova et al., 1998). Это привело к резкому возрастанию содержания биогенов в воде, несмотря на прекращение их поступления с речным стоком. В результате, к началу 1990-х гг. произошло резкое возрастание концентрации фитопланктона, и он стал, как и в остальных морях, основным продуцентом. Изменения биотопа и продукционного блока вызвали соответствующие перестройки трофической структуры и таксономического состава зоопланктона, зообентоса и ихтиофауны: из 49 видов зоопланктона сохранилось только 7, из 61 видов зообентоса — 27, из 33 видов рыб — 9 (Андреева, 1989; Андреев, 1999). Фактически за период менее 30 лет сформировалась совершенно новая экосистема. Детально были проанализированы изменения, произошедшие с Bivalvia (Андреева, 2001; Андреева, Андреев, 2001, 2003). Массовое вымирание привело к появлению и накоплению в Арале неиспользуемых ресурсов: к первой половине 1980-х гг. уцелел случайный набор видов, некоторые трофические группировки были полностью утрачены. В зоопланктоне, зообентосе и нектоне исчезли практически все хищники. В зообентосе сохранились только собирающие детритофаги и один фильтратор инфауны. Несмотря на резкое увеличение концентрации фитопланктона, потреблявшие его фильтраторы эпифауны вымерли. Для потребления неиспользуемых ресурсов симпатрически возникли новые таксоны, в первую очередь из предкового вида Cerastoderma glaucum (= C. umbonatum, C. lamarcki, C. isthmicum), перешедшего от фильтрации из прилежащего к грунту слоя воды (фильтратор инфауны) — тип питания большинства Cardiidae — к фильтрации из толщи воды (фильтратор эпифауны рыхлых грунтов) — тип питания, вообще не свойственный современным Cardiidae (в Паратетисе таксоны Cardiidae с таким типом питания возникали неоднократно). Освоение новой адаптивной зоны сопровождалось “рядом ароморфных преобразований... из наиболее существенных изменений морфологии раковины следует отметить новые варианты конфигурации створок с удлинением заднего края раковины и сильным смещением вперёд макушек, уплощение раковины, резкое уменьшение числа рёбер, изменение строения лигамента и замка. В начале 1990-х гг. сформировалось несколько групп, изменчивость между некоторыми из них превышает межвидовую изменчивость современных и ископаемых Cerastoderma, а по отдельным морфологическим признакам (замок, лигамент) выходит за пределы рода и даже семейства” (Андреев, Андреева, 2001: 42) (Рис. 1.27). Материнская форма также сохранилась. В другой из переживших перестройку групп Bivalvia — Abra — из одного материнского вида A. segmenta (= Syndosmiasegmentum), также недавнего вселенца из Средиземного моря, образовалось три формы. Эти формы поразительно соответствуют видам Hypanis, вымершим в Арале в результате перестройки (Рис. 1.28). В обоих случаях (с Cerastoderma и с Abra) новые формы устойчиво сосуществовали, что позволяет говорить об их таксономической самостоятельности. Таким образом, не более, чем за 10—15 поколений образовалось несколько видов моллюсков, с разным типом питания и столь сильно различающихся морфологически, что их (в случае с Cerastoderma) можно отнести, как минимум, к разным родам, а через пару десятков лет, при сохранении той же скорости эволюции, возможно, привело бы и к разным семействам. Ни о каком аллопатрическом видообразовании не может идти и речи: все возникшие за это десятилетие виды симпатричны. Нет оснований предполагать наличие каких-либо прекопуляционных механизмов изоляции: все они имеют наружное оплодотворение и пелагическую личинку.

Рис. 1.26. Аральское море в 2000 г., видна береговая линия середины прошлого века. По Аладину и др., 2004).
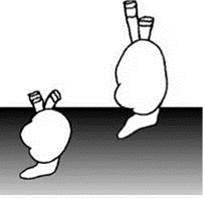
Рис. 1.27. Разные экотипы Cerastoderma Аральского моря. По С.И. Андреевой (2001).
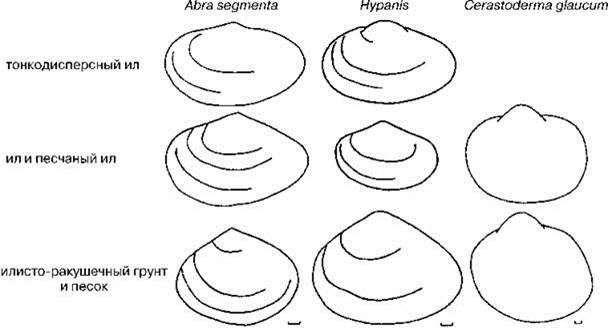
Рис. 1.28. Жизненные формы инфаунных двустворчатых моллюсков Арала. По С.И. Андреевой (2001).
К сожалению, дальнейшее формирование экосистемы Аральского моря проследить не удастся: из-за всё возраставшей солёности ныне оно превратилось в гипергалинный почти безжизненный водоём, все описанные выше виды уже вымерли. Время их существования составило, таким образом, лишь немногим более 10 лет.