Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Тропическая зона - Основные зоны - Широтная зональность - Биогеография суши
В данной книге приняты четыре зоны наземной биоты по С.М. Разумовскому: тропическая зона (17,7% от площади суши), субтропическая зона (28,0%), умеренная зона (23,5%) и бореальная зона (30,8%) (Рис. Ц4).
Тропическая зона
Резкое возрастание климатического градиента “экватор-полюс” на границе эоцена и олигоцена привело к выделению умеренного и тропического поясов из единого субтропического пояса (Рис. 3.17).
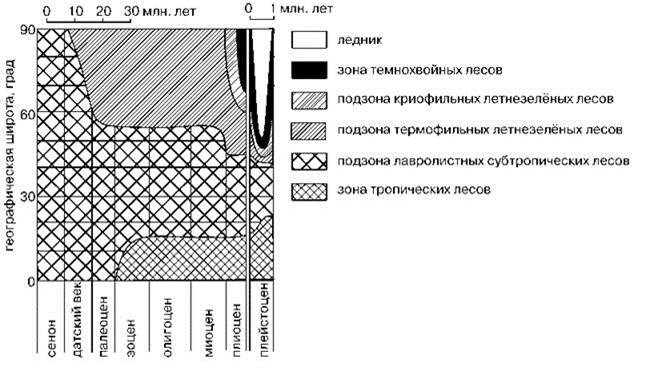
Рис. 3.17. Изменения растительности Северного полушария с конца мела (для плиоцена и плейстоцена масштаб времени увеличен). По Разумовскому (1971).
Климат зоны равномерно жаркий, поэтому сезоны года (если они имеются) определяет ход влажности, а не температуры. Климат может быть постоянно влажным или иметь сухой сезон, продолжающейся несколько месяцев.
В тропической зоне (Рис. Ц4) можно выделить два основных типа СС: лесные и мозаичные. Основной тип леса в районах с достаточным увлажнением — тропических дождевой лес (ТДЛ), районы с меньшим увлажнением заняты листопадными тропическими лесами (в лесных климаксах господствуют деревья с мелкими мезофильными листьями, одновременно опадающими в сухое время года). Очень большие площади заняты мозаичными СС (саванны). Хвойные леса (как и вообще виды хвойных) в этом поясе совершенно отсутствуют. Обширные площади, ранее занятые тропическими лесами (Рис. 3.18 и Рис. Ц4) в настоящее время заняты ранними сукцессионными стадиями (в том числе и безлесными) после выжигания леса, подсечно-огневого земледелия и просто вырубок.
Тропические дождевые леса (Pluviisilvae)
Эдификаторы лесных климаксов — деревья с вечнозелёными плоскими блестящими неопушенными листьями, без почечных чешуй. Редукция почечных чешуй легко объяснима в климатических условиях тропиков. Эдификаторами климаксных ассоциаций ТДЛ всегда являются бобовые. Несмотря на то, что термин “ТДЛ” широко распространён, разные авторы относят к ним несколько разные области. В любом случае ТДЛ расположены в приэкваториальных районах Южной и Центральной Америки, Африки, Мадагаскара, юго-востока Азии и северовостока Австралии (Рис. 3.18 и Рис. Ц4), хотя площадь, ими занимаемая варьирует от автора к автору.
“Область вечнозелёного тропического дождевого леса претерпела большие изменения под влиянием человека. В густонаселённых районах тропиков, например на Яве, не сохранилось ни одного не потревоженного человеком участка леса” (Вальтер, 1968: 189).
Общепринято считать, что для биотопа ТДЛ характерно:
— высокое количество осадков — до нескольких метров в год, хотя в Бразилии описаны редкостойные, но сходные с ТДЛ по флористическому составу и почвам леса, существующие при годовой сумме осадков около 800 мм (Santmo de Assis, 1985). Не совсем понятно, сколь велик вклад самих лесов в суммарное количество осадков, но известно, что сведение ТДЛ может уменьшать количество осадков.
— высокая температура — среднегодовая 22—28°, или средняя самого холодного месяца не ниже 18°. Такую температуру отмечают даже в тропиках только в низинах, поэтому не всякий лес, растущий в астрономических тропиках — тропический. В частности, горные леса тропиков по таксономическому составу следует считать субтропическими.
Примеры климадиаграмм районов ТДЛ приведены на Рис. 3.19. Однако климадиаграммы характеризуют климат открытых пространств и, отчасти, условия верхнего яруса. Влажность в приземном слое близка к насыщению, что исключает существенные колебания температуры. Эту влажность обеспечивают не только дожди, но и большое количество росы, конденсирующейся в кронах. Освещённость под пологом падает в 100—200 раз (что соответствует условиям под пологом широколиственных лесов умеренного пояса и достаточно для теневыносливых растений) и даже до 1000 раз (что не позволяет существовать большинству растений). Поэтому в ТДЛ почти нет подлеска и травяного покрова (число видов трав меньше числа видов деревьев), но зато широко распространены лианы (более 90% всех видов лиан растут в ТДЛ (Schrenk, 1892, 1893) и эпифиты (за пределами ТДЛ сосудистые эпифиты почти не встречаются) (Вальтер, 1968).
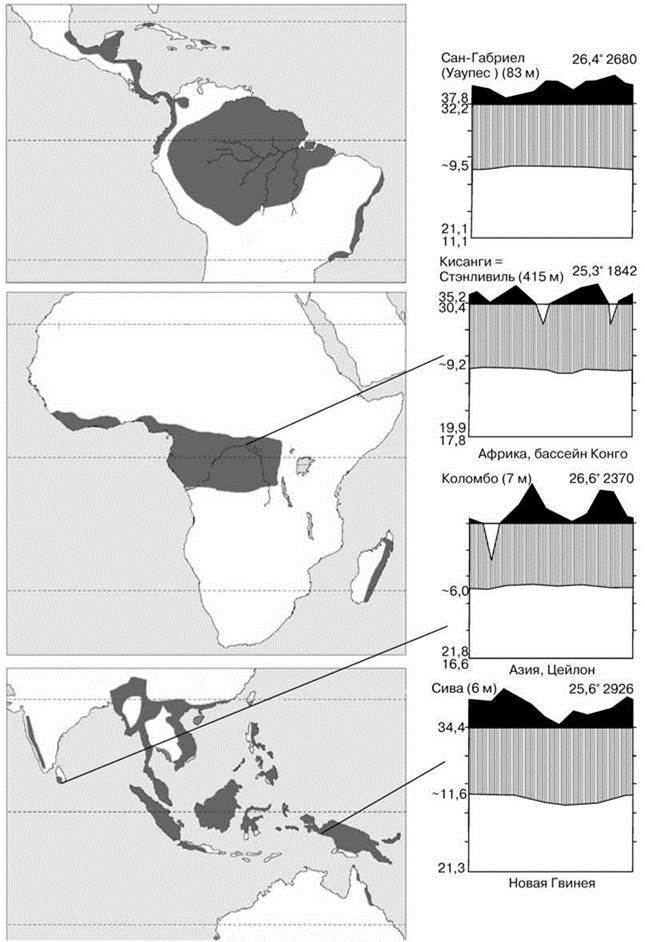
Рис. 3.18. (Слева). Восстановленный ареал тропических дождевых лесов. По Primack, Corlett (2005). В действительности горные районы покрыты субтропическими лесами (см. Рис. 3.19)
Рис. 3.19. (Справа). Примеры климадиаграмм районов, занятых тропическим дождевым лесом. По Г. Вальтеру (1968).
Внешний облик ТДЛ на всей Земле одинаков. В ТДЛ можно выделить два-три яруса деревьев и ярус подроста, кустарников и высоких трав. Стволы деревьев верхнего яруса прямые, ветвящиеся максимум 3-4 раза, против 5-6 в умеренных лесах, многие деревья совсем не ветвятся (пальмы, древовидные папоротники, некоторые двудольные), поэтому кроны их небольшие. Кора гладкая, очень светлая и тонкая, корки обычно нет. Высота деревьев 30-40 м, что соответствует таковой широколиственных лесов и тайги, но из-за высокого видового разнообразия и, следовательно, разной высоты деревьев, верхняя граница крон не прямая, как в умеренных лесах, а сильно зубчатая. Возраст деревьев косвенно (из-за полного отсутствия годовых колец) оценивают в 200-250 лет. Это значительно меньше возраста деревьев-эдификаторов лесных климаксных ассоциаций неморальных лесов (дубы) и тайги (ель, пихта). Корневая система, судя по вывороченным деревьям, поверхностная, поэтому необходимую устойчивость деревья первого яруса получают за счёт досковидных корней. Форма и размер листьев весьма однообразны, независимо от таксона: деревья в подавляющем большинстве случаев имеют большие цельные тёмно-зелёные листья с развитой кутикулой, блестящие, нередко с кончиком- капельницей. Общий их облик сходен с листьями лавра или лавровишни. Примером таких листьев является широко известный фикус, выращиваемый как комнатное растение. Молодая листва часто выглядит увядшей и окрашена в оттенки красного, в умеренных широтах подобное наблюдается редко, например, у клёна платановидного Acer platanoides и некоторых других клёнов, в меньшей степени у липы Tilia microphilla и дуба Quercus robur. По мере зеленения она становится жёсткой. Молодые листья появляются круглогодично или, если есть выраженные сезоны с разным увлажнением, то в начале сезона дождей. Несмотря на то, что климадиаграммы показывают избыток воды, сухие периоды бывают во всех экваториальных областях. Поскольку расход воды транспирирующей листвой очень велик, а неглубокая корневая система использует только часть почвенной воды, недостаток воды — обычное явление. Листья многих деревьев расположены вертикально, поэтому при высоком стоянии солнца они получают только ослабленный свет и ассимиляция в тропиках не выше, чем в других районах.
“Если вспомнить, что деревья верхних ярусов имеют нередко как бы жестяные, жесткие листья ксероморфной структуры, а травянистые растения — пластинки из одного слоя клеток, то очевидна та огромная амплитуда в условиях существования, которая имеет место в различных ярусах тропических лесов. Представление о том, что все растения влажных тропических лесов являются ярко выраженными гигрофитами, неправильно: фитоклимат здесь крайне разнообразен” (Алёхин и др., 1961).
Представление о джунглях из голливудских фильмов, в которых пробраться можно только, прорубая дорогу в густых зарослях, в действительности относится к ранним сукцессионным стадиям восстановления после вырубки (вторичные леса), опушкам и берегам водоёмов. Нижний ярус иногда вовсе отсутствует. Поскольку какой-либо (травяной, моховой или лишайниковый) напочвенный покров также отсутствует, больше неприятностей доставляет скользкая глинистая почва, а не густая растительность.
Многочисленные в ТДЛ эпифиты живут в лучших условиях освещённости, экономя на стебле. Однако одновременно они лишаются и доступа к ресурсам почвы, в первую очередь к воде. Одни, как бромелиевые, решают эту проблему путём создания водоёмов в пазухах листьев, другие стали суккулентами, т. е. имеют развитые водозапасающие ткани. Наиболее известны из них кактусы, которые многие считают свойственными исключительно пустынным ландшафтам, хотя две близкородственные трибы этого семейства (Rhipsalideae и Hylocereae) с примерно сотней видов включают почти исключительно эпифиты ТДЛ*.
Классические ТДЛ, или гилеи**, изобилующие лианами и эпифитами, представляют собой стадии гидросериальных и аллювиальных мезосериальных экогенетических рядов и особенно сильно развиты в избыточно влажных экваториальных частях зоны. В более сухих периферических частях они сосредоточены в долинах рек (галерейные леса). Обычно описывают ТДЛ нижних частей склонов гор. С увеличением высоты местности при некотором уменьшении температуры резко возрастает количество осадков, достигая максимума в поясе облаков. Соответственно увеличивается количество лиан и эпифитов, среди последних начинают преобладать папоротники и плауны. Увеличивается число древовидных папортников. На равнинах из-за уменьшения уклона местности часто ухудшается дренаж, поэтому пальмы, хорошо переносящие периодическое затопление, играют большую роль, зато лиан и эпифитов меньше. В настоящее время ТДЛ низменностей почти повсеместно сведены и замещены плантациями (сахарного тростника, риса, бананов и др.). Но даже, казалось бы, девственные ТДЛ в действительности на протяжении тысячелетий подвергались антропогенному воздействию. Например, ТДЛ Амазонки, который принято считать девственным, столь сильно модифицирован воздействием индейцев (не только растительность, но даже рельеф), что даже в доколумбово время его можно было считать “Cultural Parkland” (Heckenberger et al., 2003: 1710). Поэтому нельзя априорно утверждать, что сохранившиеся ныне участки “девственных” ТДЛ являются климаксными ассоциациями СС ТДЛ. В какой степени сильная мозаичность тропических лесов — их естественная особенность, а в какой — результат антропогенного воздействия и сукцессионного восстановления, сейчас установить практически невозможно.
Площадь ТДЛ составляет около 5—6% суши (Лесная энциклопедия, 1986) (хотя быстро сокращается в результате деятельности человека). Для ТДЛ характерно высочайшее таксономическое разнообразие, там всё ещё живёт более половины видов животных и растений Земли. Биомасса растений составляет порядка 350—700 т/га, животных — 1 т/га, т. е. та же, что и в лесах умеренного пояса Северного полушария или выше. ТДЛ — один из наиболее высокопродуктивных типов биомов (Рис. 3.20). Чистая продукция 6—50 т/га (Р/В коэффициент = 0,01—0,1). Впрочем, можно отметить, что в верхней сублиторали биомасса животных часто бывает выше на несколько порядков; биомасса же растений, хотя и достигает иногда и значительно больших величин (заросли Macrocystis у Кергелена — до 6600 т/га), обычно всё же на порядок-другой меньше. Однако, поскольку почти вся биомасса морских макрофитов создаётся за год, а ТДЛ за десятилетия и столетия, то продуктивность ассоциаций макрофитов вполне сопоставима с продукцией ТДЛ (Максимова, 2010) или превышает её один-два порядка. Таким образом, хотя ТДЛ — высокопродуктивные сообщества, величины их продукции не только не являются уникальными, но и весьма малы по сравнению с наиболее продуктивными СС (см. также Рис. 5.49).
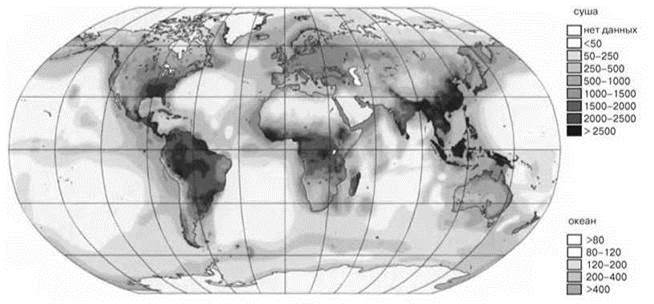
Рис. 3.20. Чистая продукция в биосфере.
Для суши и океана шкалы разные. По Berlekamp, Stegemann & Lieth http://www.usf.uni - osnabrueck.de
Высокое разнообразие животных поддерживает в первую очередь высокое разнообразие флоры: число видов одних только деревьев на 1 га ТДЛ часто превышает 100 (Walker, 1986). В результате для ТДЛ характерна исключительная сложность структуры гильдий, трофических сетей и других функциональных блоков сообществ. Сказочно сложная система ТДЛ существует в условиях крайней бедности биогенами: не менее 75% всей площади ТДЛ занято различными типами малоплодородных и неплодородных почв (Sanchez, 1989). Широко распространено мнение о том, что бедность почв — вторичное явление, что ранее почвы этих районов были гораздо более богатыми, но содержащиеся в них биогены перешли в состав организмов ТДЛ (Вальтер, 1968; Жерихин, 1993). Мнение это не подтверждено фактами. На мой взгляд, здесь более уместно совершенно другое объяснение, аналогичное с наиболее продуктивным типом морских сообществ, также отличающимся высочайшим таксономическим разнообразием — коралловыми рифами. Как ТДЛ, так и коралловые рифы существуют в условиях крайней бедности биогенами. В случае коралловых рифов эта бедность во многих случаях, несомненно, изначальная. Поэтому, вероятно, более правильно считать, что высокая продуктивность таких систем является свойством этих сообществ, сумевших накопить на бедной почве запас биогенов, высокая продукция обеспечивается высокой скоростью круговорота биогенов, а их высокое таксономическое разнообразие обеспечивает эффективность этого круговорота. Запас биогенов ТДЛ удерживает и возобновляет благодаря нескольким особенностям:
— Основные фотосинтетики — деревья. Биомасса листьев относительно невелика 1—2% всей фитомассы деревьев или 2—6% их наземной биомассы (Fittkau, Klmge, 1973;Huttel, Bernhard-Reversat, 1975; Yamakura et al, 1986), столько же, сколько в лесах умеренной зоны.
— Многие деревья, в том числе все эдификаторы лесных климаксных ассоциаций относятся к бобовым, способным фиксировать азот воздуха, способностью фиксировать азот обладают и симбиотические бактерии термитов. В умеренных широтах, хотя и есть деревья, способные фиксировать азот воздуха (например, ольха), они немногочисленны и эдификаторов климакса среди них нет.
— Переработку отмерших частей растений осуществляют преимущественно животные, а не микроорганизмы, поэтому растворимые стадии разложения практически отсутствуют. В других типах лесов консументы, как правило, потребляют менее 10% опада (Seastedt, 1984). Особенно активно его перерабатывают в дождливый сезон, когда риск потерь максимален. Так, в незатопляемых лесах Амазонии в сухой сезон половину всего опада разрушают за 218, а в дождливый — всего за 32 дня (Luizao, Schubart, 1987). Свыше половины всего объёма переработки отмерших частей растений, особенно в сомкнутом ТДЛ (Abe, Matsumoto, 1979) обеспечивают термиты Termitidae. Это семейство наиболее высокоорганизованных термитов имеет крупные долгоживущие колонии с большим (тысячи) числом особей в колонии. Termitidae резко доминируют в ТДЛ и саваннах, но слабо представлены в субтропических лесах. Их отличает от прочих семейств термитов наличие азотфиксирующих кишечных симбионтов, кроме того, в дополнение к ферментам симбиотических бактерий, они имеют собственные ферменты, разрушающие целлюлозу. Число термитников может достигать 1000 на 1 га, а биомасса термитов — 5—50 г/кв.м. Биомасса всех остальных микроартропод (коллембол, клещей орибатид и др.) суммарно на порядок меньше. Велика и роль грибов.
— Почвы и подстилка непосредственно на земле развиты гораздо хуже, чем в большинстве других наземных СС, основная часть корневых систем деревьев расположена на глубине 10—30, редко 50 см. Благодаря обилию эпифитов и корневые системы, и азотфиксаторы обильны и в надземных ярусах, где эпифиты с помощью различных приспособлений создают участки “эпифитной” почвы. Количество гумуса, содержащегося в таких “эпифитных” почвах может достигать нескольких т/га (Klinge, 1962, 1963), что вполне сопоставимо с количеством гумуса в умеренных лесах.
— Потребление минеральных веществ ТДЛ на порядок превосходит потребление бореальным. Растения потребляют примерно в 5 раз больше минеральных веществ, чем уходит со стоком (Родин, Базилевич, 1965). Речные воды экваториальной зоны слабо минерализованы — немногим более 100 мг/л. Об эффективности потребления биогенов, освобождающихся в результате разложения отмершей растительности, свидетельствует тот факт, что родниковую воду в ТДЛ можно использовать вместо дистиллированной, а электропроводность её часто даже ниже.
Одна из важнейших групп животных ТДЛ — муравьи (Greenwood, 1987). По-видимому, именно огромная численность муравьев во всех ярусах леса подавляет деятельность фитофагов и тем обеспечивает резкое доминирование детритных цепей в трофической структуре (а также эффективность листового насоса высокоствольных деревьев (Панфилов, 1961).
Высокое разнообразие покрытосеменных в ТДЛ возможно только при соответствующем наборе опылителей. Основные опылители современных ТДЛ — социальные пчёлы, способные переключаться с одних растений на другие (Roubik, 1990). В опылении участвуют и позвоночные: птицы (колибри в тропических лесах Неогеи, нектарницы в Палеотрописе, включая тропические леса юго-востока Австралии).
По палеонтологическим данным Termitidae известны с верхнего эоцена — олигоцена. Таксоны муравьёв с высокой социальной организацией и численностью семей редки до второй половины эоцена, при этом ранее не существовало группы хищных насекомоядных, сравнимой с ними по обилию и эффективности. Социальные пчёлы также достоверно известны лишь с эоцена и нет каких-либо более древних экологических аналогов. В эоцене начинается перестройка глобальной системы течений и растёт градиент экватор-полюс. С.М. Разумовский (1999) отмечает, что тропические флоры неизвестны до эоцена. Таким образом, возраст основных таксонов ТДЛ однозначно указывает на то, что их СС возникли в эоцене, вероятно, в позднем эоцене. Они гораздо моложе субтропических лесов. Напомню, что современный облик коралловые рифы приобрели значительно позже, только в миоцене-плиоцене, когда температурный градиент экватор-полюс существенно возрос и возник антарктический ледник.
В ТДЛ Африки воздух приходит из восточной Африки, по дороге потеряв существенную часть содержавшейся в нём влаги. ТДЛ Азии в значительной степени находится в области муссонного климата, с более или менее выраженным сухим сезоном. Лишь на островах Зондского архипелага, полуострове Малакка и Цейлоне сезонная динамика увлажнения не выражена. В ТДЛ Южной Америки приходит гораздо более насыщенный влагой воздух непосредственно с Атлантического океана. К тому же, пройдя над Амазонией, воздушные массы встречают на своём пути Кордильеры, поднимаются вверх, теряя влагу, стекающую полноводными реками в Амазонскую низменность. Эти особенности атмосферной циркуляции приводят не только к большей площади современных ТДЛ Неогеи, но и тому, что в сухие периоды их площадь сокращалась в значительно меньшей степени. К тому же в ТДЛ Неогеи крупных эдификаторных видов, аналогичных палеотропическим хоботным, не было: аборигенные слоны и другая мегафауна обитали лишь в мозаичных СС. Вероятно, оба этих фактора способствовали большей сомкнутости ТДЛ Неогеи, чем Палеотрописа. Это привело к тому, что среди обитателей ТДЛ Неогеи специализация пошла по пути древолазания (выработался цепкий хвост), тогда как прыгающих и планирующих форм нет. Примерами таких групп являются большинство обезьян, опоссумы, древесные муравьеды и дикобразы, цепкохвостые еноты. В Палеотрописе, наоборот, довольно много таксонов освоили планирующий полёт (лягушки, ящерицы, шерстокрылы, грызуны) или хорошо прыгают с дерева на дерево (гиббоны).