Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Новозеландская область - Схемы биогеографического районирования суши - Биогеография суши
Положение Новой Зеландии в биогеографической картине мира не вполне ясно. Во многом это объясняется длительной изоляции Новой Зеландии, приведшей к формированию весьма специфической биоты, включая мегафауну и макрофауну. Однако ранг Новозеландского региона и его подчинение не вполне ясны. Возможно более правильно рассматривать Новую Зеландию в составе Австралийской области. В то же время не все авторы сближают Новую Зеландию с Австралией. В.В. Алёхин (1944) включает её в качестве подобласти в Палеотропис. Он пишет: “Иногда [Новозеландская подобласть] причисляется к Австралийской области, но флора её в значительной большей степени носит малезийский характер... Если ниже расположенные части Новой Зеландии носят вполне малезийский характер, то её высокогорная флора имеет целый ряд черт, сходных с горами Восточной Австралии, Тасмании и даже Южной Америки” (Алёхин, 1944: 363). Вполне возможно, что Новая Зеландия, как и Индо-Малайский архипелаг не гомогенная территория, а состоит как минимум из двух биогеографических регионов.
Необходимо помнить также, что Новая Зеландия пережила на протяжении последних нескольких миллионов лет многочисленные периоды оледенения архипелага, из-за его малых размеров гораздо более катастрофичные по воздействию на биоту, чем оледенения Голарктики. Фауна позвоночных архипелага весьма специфична: тут полностью отсутствуют наземные млекопитающие, которых, впрочем, частично заменяют футлярокрылые летучие мыши Mystacinidae****, которые прекрасно бегают по земле и в поисках пищи (в основном наземных беспозвоночных) часто зарываются в опад и перегной, могут даже выкапывать норы.
Самым крупным животным были птицы моа (6 родов, включавших по последним молекулярным данным 11 видов трёх семейств, два из которых эндемичны: четыре рода и шесть видов в Emeidae, два вида Dinornis в Dinornithidae и один вид в Megalapterygidae. Размеры моа колебались от 4 м высоты и массы в четверть тонны (Dinornis robustus) (Рис. 3.52) до 25 кг (Megalapteryx didinus). Яйца достигали длины 24 см и объёма 4,3 л (= 100 куриным). Крупнейшая современная птица — африканский страус весит вдвое меньше (до 114 кг). Для моа было характерно полное отсутствие крыльев (остальные страусы их имеют) и резкий половой диморфизм, у Dinornis moa он был больше, чем у какого-либо другого вида птиц: самцы весили всего 40—90 кг, тогда как самки 80—275 кг и были в полтора раза выше (http://terranature.org/moa.htm). Моа требовалось до десяти лет, чтобы выйти из детства, ещё несколько лет — чтобы достичь половой зрелости, тогда как остальные ныне живущие птицы готовы к размножению в возрасте одного года. Общую численность всех видов моа к моменту появления в Новой Зеландии маори (XIII в.) оценивают в 158 000. Поскольку большая часть видов новозеландской биоты имеет очень небольшие размеры (хотя к моменту появления маори крупных видов было больше), охота на моа была основным источником белка для маори. Маори потребовалось менее 100 лет, чтобы почти полностью истребить все виды моа. Последние представители моа (Megalapteryx didinus) были убиты примерно к XVI—XVIII вв. Процесс уничтожения моа был ускорен сбором их яиц. Пока моа было много, полинезийцы ели лишь бёдра птиц, выбрасывая остальное, позднее, вероятно на фоне уменьшения численности моа, маори стали есть и остальные части и дробить кости, доставая костный мозг (Anderson, 1984). После уничтожения моа на фоне роста населения в диете людей Новой Зеландии возник дефицит белка и маори от охоты и рыболовства перешли к каннибализму. Кроме моа в Новой Зеландии до появления маори обитало (некоторые сохранились на грани вымирания) несколько других крупных видов нелетающих птиц: гуси (до 1 м высотой и 15 кг весом), киви (несколько родов), попугаи, крупнейший (до 12 кг) пеликан и крупная нелетающая утка. Жил здесь возможно единственный нелетающий род воробьиных Pachyplichas (2 вида).
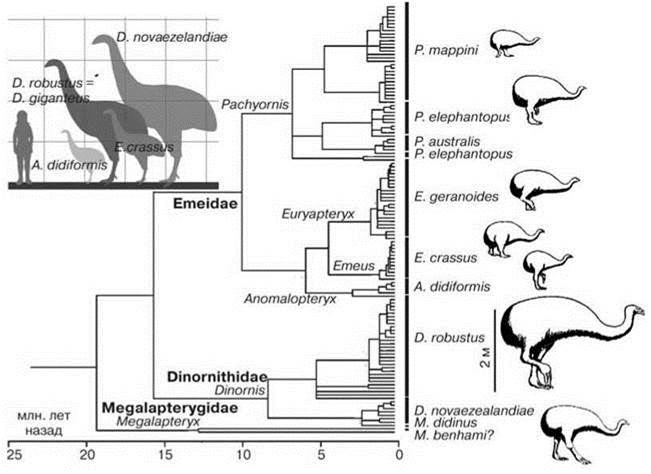
Рис. 3.52. Филогенетическое древо моа (Baker et al., 2005).
Дерево изображено в виде хронограммы. В левом углу показаны относительные размеры некоторых видов.
Крупнейшими хищниками были также птицы*****. Орёл Хааста (Harpagornis moorei) имел размах крыльев до 3 м и массу до 15 кг. Оставшись без своей привычной пищи (моа) и преследуемый маори как потенциально опасный для человека вымер примерно к 1500 г. н. э.
Другим крупными хищными птицами были два вида Aptornis — нелетающих птиц, размером с маленьких моа, высотой 80 см, весом до 19 кг.
Фауна рептилий и амфибий очень бедная, но при этом включает очень древних реликтов. Амфибии представлены только родом лягушек Leiopelma (2—3 вида), относящемуся к наиболее примитивным из ныне живущих бесхвостых земноводных Leiopelmatidae (второй род семейства обитает в умеренной зоне западного побережья Северной Америки). Leiopelmatidae выделяют в отдельный подотряд, ископаемые остатки которых известны с верхней юры. Пресмыкающиеся более разнообразны — восемь видов, в том числе хрестоматийная гаттерия Sphenodon, относящаяся к особому отряду, немногим более древнему, чем Leiopelmatidae (известен с раннего мезозоя).
Насекомые высокоэндемичны, при этом многие широкораспространённые группы малочисленны (например, дневные бабочки, бражники, прямокрылые, термиты, златки, скарабеи, скакуны) или вовсе отсутствуют (бронзовки). Нет в Новой Зеландии и скорпионов. В Новой Зеландии сформировалась разнообразная фауна (около 70 видов) гигантских нелетающих кузнечиков — вета (weta), которые взяли на себя экологическую роль специализированных распространителей семян растений с сочными плодами, которую в остальных регионах играют млекопитающие. Кузнечики охотно поедают разнообразные плоды, и часть семян при этом действительно проходит через пищеварительный тракт неповреждёнными, их всхожесть даже немного улучшается (Duthie, Gibbs, Burns, 2006).
173 эндемичных вида дождевых червей имеют ближайших родственников на юге Южной Америки, Фолклендах, юге Африки, Новой Каледонии и Австралии. Spenceriellagigantea достигает длины 1,4 м и ширины 11 мм и люменисцирует столь ярко, что можно читать в её свете (http://terranature.Org/weta.htm#earthworms).
Зато флора, хотя и высокоэндемична (порядка 50%) на видовом уровне, на родовом уровне весьма сходна с Австралийской: 85% родов тайнобрачных и 75% родов покрытосеменных общи для Австралии и Новой Зеландии. Сходство на родовом уровне с Южной Америкой вдвое ниже: 40% тайнобрачных и 43% покрытосеменных общи для Новой Зеландии и Чили.
Ареал некоторых таксонов охватывает Зондский архипелаг, включая Филиппины, Новую Гвинею, тропические леса северо-востока Австралии, и прилежащие острова (Новую Каледонию и др.). Таков ареал Agathis, входящего в семейство Araucariaceae. Ареал других родов этого семейства (включающего всего три рода) включает Новую Гвинею, Новую Каледонию и восточную Австралию и Южную Америку. Возможно, уничтоженные человеком хвойные леса Австралии образовывали араукарии. Одна из крупнейших новозеландских араукарий — каури Agathis australis — достигала 5 м в диаметре.
До заселения человеком Новая Зеландия была покрыта широколиственными лесами с лианами и папортниками, в том числе и древовидными. Эдификатор климакса — южный бук Nothofagus. Туссоковые злаковники Новой Зеландии имеют антропогенное происхождение и возникли в результате выжигания леса маори (McWethy et al., 2010;Bowman, Haberle, 2010).