Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Районирование Голарктики - Голарктика - Схемы биогеографического районирования суши - Биогеография суши
Южная граница Голарктики является северной границей Палеотрописа (в Евразии) и Неогеи (в Северной Америке). В Африке границей принято считать Сахару. В действительности по фауне млекопитающих и птиц Сахара ближе к более южным районам Африки, тогда как по фауне насекомых и наземных моллюсков — к Голарктике. Как бы то ни было, учитывая недавнее и по всей вероятности антропогенное происхождение этой пустыни, следует признать, что СС Сахары были разрушены, а новые не возникли из-за крайне неблагоприятных условий и короткого времени существования Сахары. Территория эта, таким образом, является переходной областью неопределённой принадлежности.
В СС Голарктики после последнего ледникового периода произошло массовое выпадение крупных млекопитающих (Рис. 3.58), птиц и рептилий. В северной Голарктике уничтожение мамонта привело к исчезновения “слоновьего” ландшафта. Подробно “слоновий ландшафт” рассмотрен в начале этой Главы, здесь лишь можно напомнить, что африканские слоны деревья толщиной до 35 см (редко до 70 см) выворачивают с корнями; у более крупных обдирают кору — фактически почти любое дерево Голарктики хоботные могли вывернуть с корнями. Пришедшая на смену мозаичности поясность привела к распространению на огромных площадях одних ассоциаций, тогда как другие ассоциации на тех же площадях стали редкими или вовсе исчезли. В результате широко распространились диаспорические субклимаксы.
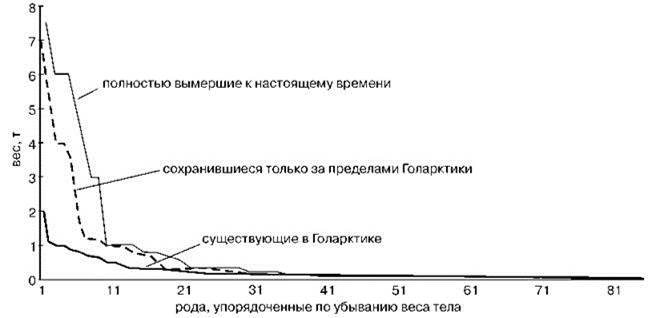
Рис. 3.58. Распределение по размеру родов крупных млекопитающих Голарктики, существовавших в течение последних 15000 лет по Жегалло и др. (2001).
Исчезновение “слоновьего” ландшафта в умеренном климате имело и другие эффекты, отсутствующие в тропических и субтропических районах:
Существующий на протяжении нескольких месяцев снежный покров нередко весьма глубок. Ныне глубокий снег и гололёд — одна из основных причин гибели копытных, происходящей зачастую при отсутствии недостатка корма. Частично в охотничьих хозяйствах проблема решается прокладкой дорог, однако сеть их обычно недостаточна (Данилкин, 2008). Мигрирующие стада мамонтов создавали гораздо более густую сеть троп, что улучшало условия обитания остальной мегафауны.
Другая причина массовой гибели копытных зимой — отсутствие водопоев, поскольку водоёмы зимой покрыты льдом. Нередко вскрытие трупов показывало, что их желудки заполнены спрессованным кормом, тогда как кишечники пусты, что свидетельствует о недостатке воды (Данилкин, 2008). И в этом случае стада мамонтов, которые также нуждались в воде, решая проблему водопоев для себя, решали её и для других, подобно тому, как в пустынных районах Африки слоновьими колодцами пользуются и другие животные.
Ниже описаны виды, вымершие в Евразии и Северной Америке, и даны краткие характеристики их биологии и роли в СС.
Евразия
В северную Евразию человек (Homo georgicus) проник не позднее 1,8 млн. лет назад. Этим возрастом датированы его находки на территории Грузии. Наиболее древние находки Homo в Европе датированы 1 млн. лет назад (Carbonell, 2008). Некоторые виды животных, вымиранию которых, как минимум, способствовал человек изображены на Рис. 3.59.
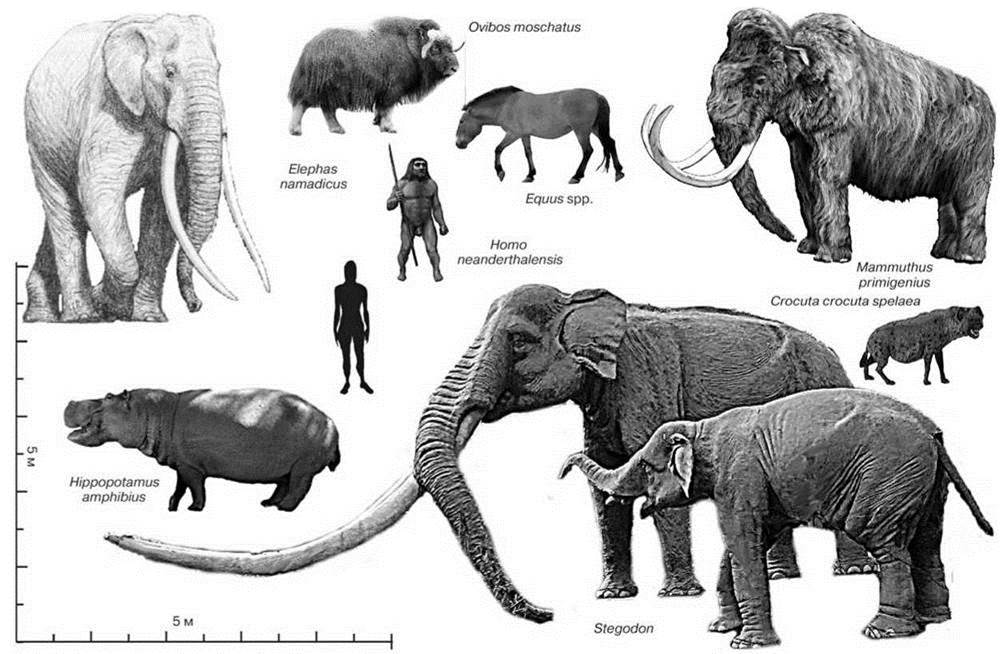
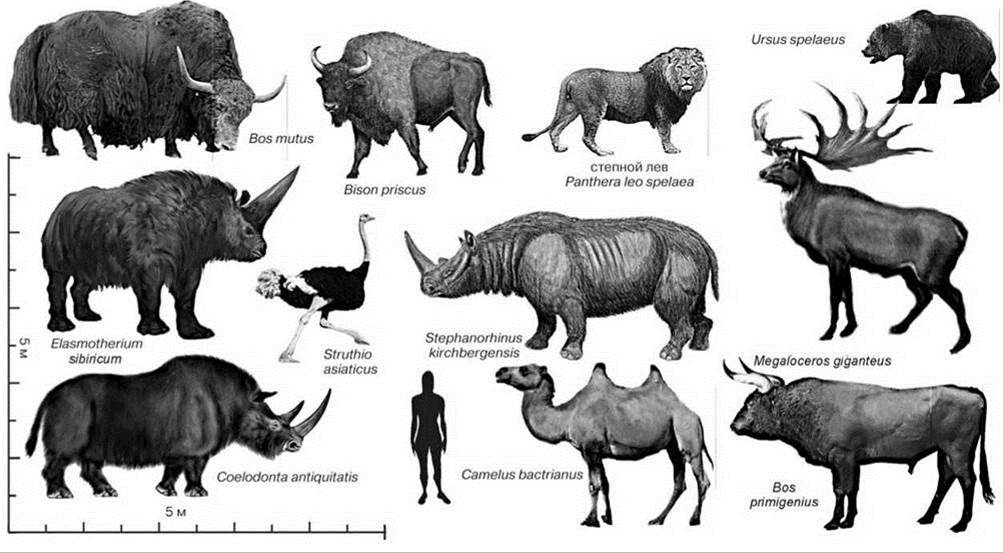
Рис. 3.59. Некоторые представители мегафауны, полностью или почти полностью исчезнувшие с появлением Homo spp. в Палеарктике (не показаны многочисленные виды, лишь заходившие в южные части Палеарктики из тропической Азии.
Хоботные. Таксономия хоботных довольно запутана, поскольку по большей части сохранились только зубы. Были представлены несколькими родами, помимо ныне живущих индийского Elephas (иногда из него выделяют отдельный род лесной слон Palaeoloxodon), мамонт Mammuthus, Phanagoroloxodon, кроме этого, на юг Дальнего Востока из тропиков Азии заходил Stegodon, а в Малую Азию африканский слон Loxodonta.
Шерстистый (woolly) мамонт Mammuthus primigenius — наиболее известный вид рода. Высота до 2,75 м. Наиболее южные находки — Ливан, северные районы Казахстана, север Монголии, Китай до среднего течения Янцзы, Сахалин и Хоккайдо. В южных частях своего ареала мамонт ходил по рекам, в которых плавали крокодилы и аллигаторы. На севере распространение мамонта ограничивала береговая линия. Фактически ареал мамонта охватывал всю территорию Голарктики (Рис. 3.60).
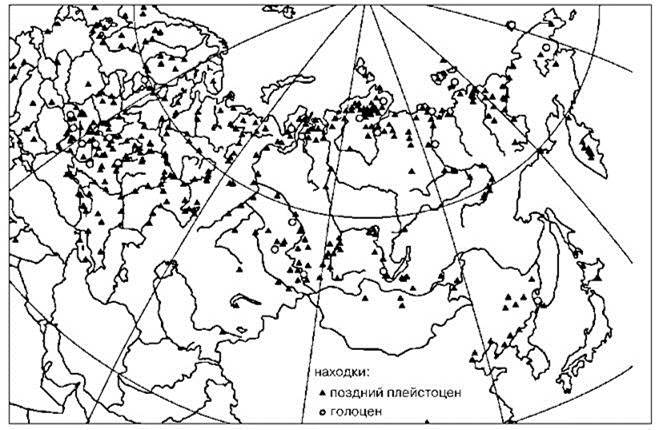
Рис. 3.60. Палеарктическая часть ареала мамонта в позднем плейстоцене и голоцене. По: Калякин, Турубанова (2004).
Тем не менее, мамонта почему-то считают видом, связанным с ледником. Возможно, с этим заблуждением связано широкое распространение мнения о вымирании мамонта в связи с потеплением климата: ледник растаял, произошло изменение экосистем, в частности тундро-степь сменили леса. Рассмотрим более подробно эту аргументацию. Климатические изменения последних 400 000 лет, можно грубо схематично представить, как два крупных ледниковых периода (рис и вюрм), между которыми находился теплый период — рисс-вюрм, во время которого климат Евразии был значительно теплее сегодняшнего. На Русской равнине тогда вместо ельников и березняков росли буковые леса. Считают, что среднегодовая температура в средних широтах была выше современной примерно на 2°С (Agusti, Anton, 2002). Мамонтовая фауна в этот период ничуть не пострадала. Мамонты входили в состав самых разнообразных СС, на севере и в средней полосе Евразии и Северной Америки это были лесо-тундро-степи, южнее — криофитные саванны. Вид был эдификатором таёжных и неморальных СС. Анализ митохондриальной ДНК сибирских мамонтов показал, что все они представляли собой единую большую семью, не подразделённую на какие-то отдельные кланы или субпопуляции. Современные индийские и африканские слоны гораздо более полиморфны (Rogaev и др., 2006). Это означает, что в течение года мамонты совершали миграции через всю Сибирь. Правомочно предположить, что способность к ежегодным миграциям на тысячи километров была присуща мамонтам и других районов Голарктики. Поэтому выяснение “климатической ниши” мамонта по местам его находок (Nogues-Bravo et al., 2008) неправомочно: в разные сезоны года мамонты обитали в разных климатических зонах, выбирая в каждый конкретный момент подходящие условия. Невозможно представить, как должен был измениться климат по всей территории Евразии и Северной Америки, чтобы он стал непригоден для такого эврибионтного вида, который пережил более сильные климатические изменения в прошлом. Если климат где-то стал для него неподходящим, мамонт просто мог туда не ходить. Палеоклиматические данные свидетельствуют о том, что и в периоды межледниковий и в периоды оледенений условия на огромных площадях Евразии и Северной Америки были вполне подходящими для мамонта. Тем не менее, в древнем и раннем голоцене мамонт исчезает почти по всей территории Северной Евразии. При этом каких-либо закономерных изменений границ ареала по сравнению с поздним плейстоценом не отмечено. Остатки костей мамонта найдены в тех же районах, где он обитал в конце плейстоцена. Анализ 310 датировок из более, чем 140 местонахождений, проведённый в Институте геологии СО РАН, показал, что популяция мамонтов Сибири вплоть до 12 тысяч лет назад успешно переживала неоднократные колебания климата и соответствующее им изменение растительных зон. Лишь около 12 тыс. л.н. началось необратимое сокращение ареала мамонтов. Наиболее поздние находки мамонтов сделаны на изолированных островах: на острове Врангеля, где он обитал ещё около 4000 лет назад и на острове Св. Павла (один из островов Прибылова) на юго-востоке Берингова моря, где он вымер примерно в то же время. Островные популяции мамонтов были гораздо мельче материковых. На о. Врангеля мамонты были всего 1,8 м высотой, причем уменьшение высоты на 1 м (>40%) произошло за 5000 лет (около 100 поколений мамонтов) (Sukumar, 2003). Выживание мамонтов на этих островах в течение многих тысячелетий после окончания ледникового периода и мгновенное исчезновение после появления на них человека является дополнительным подтверждением справедливости теории blitzkrieg. Выпадение из экосистем мамонтов не могло не оказать влияние на сами экосистемы. Вряд ли можно сомневаться в том, что, как и в хорошо документированных случаях по африканским экосистемам (см. выше), распространение лесов является следствием, а не причиной вымирания мамонта.
Хотя мамонт из-за плотного телосложения весил гораздо больше современных слонов, размером он уступал африканскому слону.
Лесной (прямобивневый, древний) слон Elephas antiquus (в Европе) и E. namadicus и (в Азии) экологически замещал мамонта в бореальных и субтропических лесах от Британии до Северного Китая. Вымер на большей части ареала 30000 лет назад. Последними сохранились популяции мелких южных слоников (ростом менее 1 м) на островах Средиземного моря (Кипре, Сицилии, Мальте и некоторых других), их обычно выделяют в отдельный вид Elephas falconeri или в несколько видов, эндемичных для каждого острова (Рис. 3.61). Эти популяции перебил, вероятно, уже Homo sapiens.
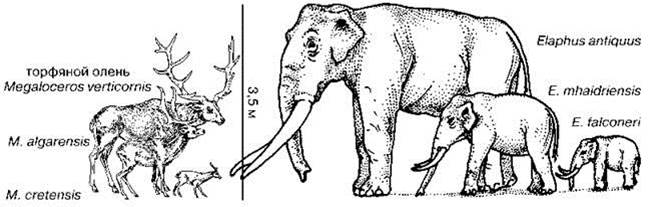
Рис. 3.61. Материковые и островные формы торфяного оленя и южного слона. По: Brown, Lomolino (1998).
Phanagoroloxodon irtyshensis — находки этого вида единичны и ограничены югом западно-сибирской низменности (Shpansky, 2005; Шпанский, Печерская, 2009). Не известно, причастен ли человек к его вымиранию, но, судя по возрасту находок, это вероятно.
Шерстистый носорог Coelodonta antiquitatis населял всю голарктическую Евразию (от Британских островов до Камчатки), но отсутствовал, как и все прочие носороги, в Северной Америке, вероятно из-за более осёдлого, чем у мамонта, образа жизни. В каждый момент времени обитал севернее остальных палеарктических носорогов. Наскальные изображения шерстистого носорога многочислены, но за исключением ангарских стоянок, нет свидетельств того, что он был объектом активной охоты, хотя в кухонных остатках встречается постоянно. Шерстистый носорог, как, вероятно, многие другие виды, получившие широкое распространение во время ледниковых периодов, сформировался на Тибете (Tao Deng et al., 2011). Вес до 4,0 т; высота в холке до 2,0 м; длина тела до 3,6 м, хвоста до 0,6 м. Хотя его называют шерстистым, шерсть, судя по найденным замерзшим трупам, не превышала по длине шерсть современных зубров, поэтому обитание вида в каком-то месте не может быть показателем “ледниковых” условий. Исчезли шерстистые носороги уже в голоцене.
Эласмотерии Elasmotherium. С территории Западной Европы эласмотерии не известны, на востоке доходили до Китая (E. peii) или Средней Азии и юга Сибири (E. sibiricus) Был гораздо крупнее современных носорогов (вес до 6,0 т; высота в холке до 2,3 м; длина тела до 4,3 м). Легко отличимы от остальных носорогов одним, а не двумя рогами. Изображён на наскальных рисунках. Многие считают, что именно он послужил прообразом сказочного единорога. Судя по строению зубов обитал в окрестностях пресных водоёмов, возможно вёл полуводный образ жизни. Позднейшие находки (Южное Зауралье) 40—20 тысяч лет назад.
Носороги Мерка Stephanorhinus. S. kirchbergensis (от Европы до Китая, в том числе и в северной Азии, вплоть до Вилюя, а, возможно, и дальше), предпочитавший леса и S. hemiotechus, предпочитавший более открытые ландшафты и не проникавший в северную Азию. Носороги весом до 3 т, высотой до 2 м и до 4 м длиной. Строение скелета позволяет предположить, что они были хорошими бегунами.
Разнообразны были олени. Самый крупный вид — гигантский (большерогий, торфяной, ирландский) олень Megalocerous giganteus. Достигал 2,1 м в плечах и веса в 700 кг. Размах рогов этого вида, достигавший 3,7 м свидетельствует о том, что он обитал в открытых и полуоткрытых ландшафтах. Рога этого оленя часто попадались в европейских торфяниках и хозяева средневековых замков вывешивали их на стены, создавая впечатление, что это они их убили. Ареал вида простирался от Ирландии до Байкала. На Южном Урале жил ещё 7 тысяч лет назад (Stuart et al., 2004).
Лоси Alces. Предпочитают мозаичные ландшафты, современная их приуроченность к лесам вторична. В плейстоцене лоси были немногочисленны, резкое возрастание поголовья в Евразии произошло, видимо, в раннем голоцене (Данилкин, 2008). Современные лоси относятся к двум видам, различающимся числом хромосом и морфологическими признаками: европейский лось A. alces и американский лось A. americanus. Ареал последнего охватывает не только Северную Америку, но и Восточную Сибирь и Дальний Восток. Виды, по-видимому, способны к скрещиванию (Боескоров, 2001).
Сайга Saiga tatarica. В конце плейстоцена — начале голоцена от Атлантики до Аляски и Канады, на север до Новосибирских островов. Вид — хороший индикатор малоснежных, а вовсе не степных районов (Сайгак..., 1998).
Северный олень Rangifer tarandus. Этот вид, как и мамонта, почему-то считают характерным для тундры. Выше уже указано, что тундра — это часть зоны тайги, занятой ранними безлесными стадиями, большей частью гидросериальными. Отсутствие древесных стадий сукцессий, вероятно, обусловлено антропогенным воздействием. Северный олень — вид, свойственным этим, а также пирогенным ксеросериальным стадиям и аналогичным стадиям сукцессий неморальных лесов. Ещё в XVIII в. в периоды зимних кочёвок он регулярно заходил на Украину, север Калмыкии и постоянно обитал в Башкирии, в XIX в. — в Ленинградской, Костромской и Горьковской областях, где до сих по находят их сброшенные рога. На юго-востоке Тверской области окончательно уничтожен в начале XX в. До настоящего времени сохранился в Монголии, северо-западе Маньчжурии, Амурском крае, севере Сихоте-Алиня, т. е. районах, никогда и никем не относимых к “зоне тундры”. Это широко и быстро мигрирующий зверь, голоценовая форма которого адаптирована к питанию прежде всего мягкими кормами (лишайниками), а не злаками. Олени уходят с пастбищ, на которых становится мало лишайников. У северных оленей очень большая площадь копыт, малый вес и мало удельное давление на грунт, поэтому они, в отличие от лошадей и бизонов, не оказывают серьезного разрушающего воздействия на моховой покров и не вызывает его смену травяным.
Тур Bos primigenius — дикий бык из Европы и Азии, ставший прародителем большинства пород крупного рогатого скота. Это мощное животное (вес до 2 т) чёрной окраски со светлой полосой на спине полностью исчезло к 1627 г.
Бизон Bison priscus. Ареал первобытного бизона также охватывал всю Голарктику (если рассматривать североамериканского бизона как подвид) или, по крайней мере, всю голарктическую Евразию от Пиренеев, Апеннин, Босфора, на юге — Малая Азия, Кавказ, Прикаспийская низменность, предгорья Средней Азии, Монголия, север и северо-восток Китая, юг Приморья, Япония. В Европе исчез 10—8 тысяч лет назад, но на востоке Сибири (Приангарье) дожил до VIII—X вв. Обладал широкой географической изменчивостью, описано много подвидов. Расстояние между кончиками рогов достигало 2 м (Зубр, 1979).
Овцебык Ovibos. Известны изображения овцебыков на серебряных пластинках, найденных в курганах близ Улан-Батора возрастом 2 000 лет. На святилище в Ненецкой тундре был найден череп овцебыка. Судя по его сохранности, череп едва ли мог быть положен на святилище до 30-х гг. XX в. (Жегалло и др., 2001).
Лошади Equus. Широкопалая лошадь E. latipes (Русская равнина, Молдавия), уральская лошадь E. uralensis (Урал от севера до Каспия), лошадь Пржевальского E. przewalskii (Азия до Жёлтого моря), тарпан E. gmelini (Польша — южный Урал, истреблён в 1814 г.), европейский плейстоценовый осёл E. hydruntinus (юг Западной Европы — предгорья Средней Азии), кулан E. hemionus (Причерноморье — Колыма — северо-восток Китая, Аравийский полуостров).
Як. В настоящее время род представлен двумя видами — домашний як Bos grunniens и дикий як B. mutus (International Commission..., 2003). Анализ ДНК свидетельствует о близости яков и бизонов. Дикий як обитает в безлесных нагорьях от высоты минимум 3,5 км до 5,5 км над уровнем моря, широко кочуя по альпийским пастбищам и ледяным пустыням Тибета, питаясь мхом, лишайниками и травой. Способен спокойно переносить морозы до -40°С и многочасовые горные бури зимой и летом, однако практически не переносит высокую температуру и на два летних месяца уходит к вершинам выше уровня снега. В качестве адаптаций к горным биомам обладает особыми свойствами крови с эритроцитами, значительно меньшими коровьих, и, как минимум, в три раза более многочисленными. Пищеварительный тракт яка ферментирует растительную массу при высокой температуре с большим количеством выделяемого тепла и в зимние месяцы работает как внутренняя грелка. Размеры диких яков до 2,2 м в плечах, длина до 3,3 м и до 1 т весом. Самки в три раза легче. Длина рогов самки дикого яка — 50 см. Самца — более 1 метра. Домашняя форма более чем в два раза меньше дикой. Продолжительность жизни ~ 25 лет, половая зрелость примерно в 6 лет, рождение теленка 1 раз в два года, при плохих кормовых условиях, на бедных пустынных или каменных горных пастбищах реже, 1 раз в три года. Ныне стада по 20—30 голов, изредка могут собираться по 200 особей, но ещё в XIX в. по наблюдениям Н.М. Пржевальского стада достигали численности нескольких сотен и даже тысяч голов. Взрослые самцы диких яков живут либо стадами холостяков, либо по одиночке. В плейстоцене на территории Сибири был широко распространён другой вид этого рода — байкальский як (P. baicalensis). В плейстоцене як (P. mutus) был широко распространен в Северной Азии (Жегалло и др., 2001).
Бегемот Hippopotamus amphibius. Высота в плечах до 1,65 м, длина тела до 5,4 м, вес до 4,5 т. Как и слоны и копытные, бегемоты, обитавшие на островах Средиземного моря, были гораздо мельче, их часто выделяют в отдельные виды: карликовые гиппопотамы Кипра H. minor (200 кг, длина 121, высота 76 см); Сицилии H. pentlandi (до 320 кг); Крита H. creutzburgi; Мальты H. melitensis. Бегемоты в Европе исчезли во время ледниковых периодов, какую роль в их исчезновении сыграли неандертальцы, установить сложно, несомненно, однако, что карликовые виды бегемотов, эндемичные островам Средиземного моря, были истреблены человеком. Необходимо отметить, что представление о бегемотах как чисто тропических животных основано исключительно на их современном ареале, являющимся лишь небольшой частью их ареала до возникновения человека, ранее бегемоты были широко распространены по всему Палеотропису и югу евразийской части Голарктики. Бегемот в Ленинградском зоопарке пережил блокаду во время Второй мировой войны, питаясь опилками.
Африканский страус Struthio camelus. Во Внутренней Монголии африканский страус, возможно, встречался в II—I в д.н.э. (Карташев, 1974), так что разведение страусов в средней полосе России — это не причуда, а реинтродукция.
Саблезубые кошки. В плейстоцене на севере Евразии обитал Homotherium latidens (размером со льва или тигра, до 1,2 м в плечах, вес до 350 кг).
Лев Panthera leo. Ареал льва в начале голоцена охватывал всю Африку, Евразию и Северную Америку. В историческое время ареал распространения львов охватывал всю Африку, Балканский полуостров и Армению, Аравийский полуостров, Турцию, Ирак, Сирию, Иран, Афганистан, Индию. В Греции львы исчезли к концу I в. н. э., в Крыму — в V—II вв., в Закавказье — в Х в. н. э., в Иране последний лев был убит в 1930 г. (Соколов, 1979, 1990).
Тигр Panthera tigris. Останки тигра (не пещерного льва) обнаружены на пространстве от центральных районов Якутии до Юго-Восточной Азии, включая весь Дальний Восток и Японские острова (мелкая форма), на запад до Ирана и Кавказа. Тигры на Кавказе до начала XIX в. тоже были, но витязь в тигровой шкуре появился из-за безграмотности переводчика: в оригинале шкура была леопарда.
Леопард Pantherapardus. Ареал леопарда захватывал почти всю Европу, реликтовые популяции оставались в Крыму довольно долго. На Кавказе обитал в историческое время.
Пещерный медведь Ursusspelaeusи другие виды. Длина тела до 3,5 м, вес до 0,5 т.
Неандерталец Homo neanderthalensis. Согласно одной из точек зрения неандертальцы — потомки Каина, тогда как кроманьонцы H. sapiens — потомки Сифа. Это объясняет внешние различия между древними людьми как проклятье Божье каинитов (Быт. 4:11), утратившим красоту первозданного Адама, а также как действие “знамения”, которое Господь “сделал” Каину, чтобы никто его не убил (Быт. 4:15) (Волков, 2008). Согласно другому мнению, H. neanderthalensis отделились от H. sapiens 350—400 тысяч лет назад. Вид населял Европу и Азию на восток примерно до Байкала. Исчез, вероятнее всего в результате вытеснения H. sapiens. На стоянках кроманьонцев были обнаружены обглоданные кости неандертальцев со следами зубов, характерными царапинами и разломами на костях. H. sapiens делали ожерелья из зубов неандертальцев. Наиболее поздние находки неандертальцев датируют 24 000 лет назад (на территории Пиринейского полуострова).
Ещё одним важным компонентом экосистем Голарктики являются бобры. Их ареал охватывал всю северную Евразию, включая северный Китай (Лавров, 1981). Семейство эндемично для Голарктики, в Евразии один вид. Весьма вероятно, что сдвигание на север границы ели и пихты в значительной степени обусловлено редукцией гидросериальных рядов СС юга из-за уничтожения бобров (об эдификаторной роли бобров см. Главу 4).
Вымирание мегафауны в евразийской части Голарктики произошло не одновременно. Численность пещерного медведя резко падает одновременно с вымиранием неандертальцев в Европе и на Кавказе задолго до начала общего кризиса мегафауны — однако примерно совпадает хронологически с экспансией нашего вида. Одновременно с этим начинается угасание лесного слона и бегемотов в Европе (около 30 тысяч лет назад — некий рубеж вымирания Elephas, бегемотов, носорогов Мерка, европейских саблезубых кошек, почти полного вымирания неандертальцев и падения числа пещерных медведей). Эти события, в отличие от крушения мегафауны на границы голоценового потепления, мало связаны с изменениями климата, да и как-то странно связывать вымирание теплолюбивых видов с потеплением. Зато их вымирание совпадает с ростом численности Homo. Шерстистый носорог в Европе вымирает практически одновременно с мамонтом.
Столь стремительного вымирания, как это было в Северной Америке, в Старом Свете не было, поскольку в качестве эдификаторов “клочковатых” ландшафтов в Па- леотрописе и Голарктике хоботных могут до некоторой степени заменять носороги, крупные быки и бегемоты. К несчастью, в Северной Америке бегемотов не было вовсе, носороги хотя и возникли там, но к нижнему плиоцену уже вымерли, а крупные жвачные были представлены только одним родом (Bison), тогда как в голарктической Евразии, кроме двух видов Bison (зубра и бизона), были тур, а в Сибири и байкальский як. Южноамериканские по происхождению таксоны мегафауны: наземные ленивцы и, особенно, броненосцы, были беззащитны перед человеком, и, по-видимому, были истреблены столь же стремительно, как и черепахи. Более разнообразный состав мегафауны голарктической Евразии и более растянутое во времени воздействие человека на неё привёло к меньшему её вымиранию. Кроме того, эволюция мегафауны Старого Света происходила в значительной степени совместно с человеком, что сделало её менее уязвимой к его воздействию, поскольку давало время для модификации поведения со скоростью, сопоставимой со скоростью изобретением человеком новых способов охоты. Фактически в голарктической Евразии исчезли только хоботные и носороги, да несколько видов крупных копытных, например большерогий олень. Остальные виды или их близкие родственники до сих пор уцелели в рефугиумах или в одомашненном состоянии. Т. е. выпадение их из СС было постепенным, длилось десятки тысяч лет, что дало возможность биоте хоть как-то адаптироваться.
Северная Америка
Механизм катастрофического вымирания североамериканской биоты в результате вселения человека был рассмотрен в общих чертах в начале главы. Обратимся теперь к деталям процесса. Охарактеризуем мегафауну континента 12000 лет назад, т.е. непосредственно перед появлением человека (Рис. 3.62).
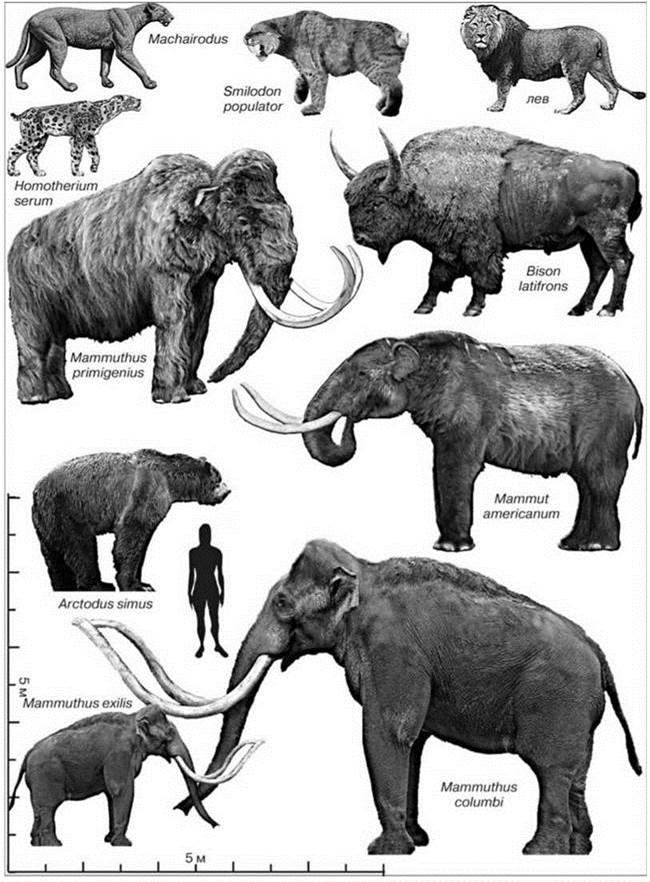

Рис. 3.62. Некоторые представители мегафауны, исчезнувшие с появлением человека в североамериканской части Голарктики.
Большая часть вымерших животных — травоядные, остальные виды — питавшиеся ими хищники или поедавшие их трупы падальщики. В современных сукцессионных системах им или нет аналогов (мамонты, гиганские ленивцы, саблезубые кошки, хищные птицы или рептилии) или их заменили более мелкие виды (гиганский бизон на бизона, тераторн на кондоров). На мелких островах крупные травоядные были представлены мелкими формами (по: Brown, Lomolino (1998), Wikipedia).
В Северной Америке (исключая её юг, относящийся к Неогее) ещё несколько тысяч лет назад обитало три основных вида хоботных. Шерстистый мамонт Mammuthusprimigenius (тот же вид, что и в Евразии) на севере, южнее обитал колумбийский мамонт (M. columbi) высотой до 4,3 м и бивнями до 5 м длиной и весом почти до 10 т. Мамонты доходили на юг до центральной Америки. С ними сосуществовал (за исключением Аляски) представитель другого семейства — американский мастодонт (Mammut americanus) высотой до 2,75 м. Мастодонты в отличие от мамонтов, имели зубы с коническими вершинами или гребнями, а не со складчатой эмалью. Такие зубы были годны для пережевывания относительно мягких побегов, хвои и листвы деревьев. Ещё южнее, в тропиках Неогеи (включая Флориду) обитал другой род мастодонтов — Cuvieronius, заходивший сюда из Южной Америки. На островах Алеутского архипелага и Channel Islands на севере Калифорнии обитали островные популяции M. primigenius и M. columbi, соответственно. Как и на островах Средиземного моря, особи островных популяций были гораздо мельче материковых, например островной M. columbi, выделяемый в отдельный вид M. exilis, был всего 1,72 м высоты при весе в 760 кг. Островные популяции сохранялись гораздо дольше материковых, но также были истреблены людьми, как только они попали на острова. Так мамонты на острове Святого Павла (Алеутская гряда) были истреблены только к 4000 г. до нашей эры.
В состав СС Северной Америки входили и южноамериканские по происхождению таксоны: два семейства гигантских броненосцев весом до 1 т (Glyptodontidae иPampatheriidae) и четыре семейства гигантских ленивцев. Eremotherium laurillardi (Megatheriidae) — один из самых крупных ленивцев, — достигал длины 6 м и веса более 3 т. Nothrotheriops shastensis (Nothrotheriidae) — один из самых маленьких ленивцев — всего в 250 кг веса, при длине 2,75 м от кончика морды до кончика хвоста, его субфоссильный помёт находят в пещерах (к этом же семейству принадлежал упомянутый выше южноамериканский Thalassocnus natans, питавшийся морскими макрофитами). Paramylodon (Mylodontidae) достигал 3 м высоты и веса более 1 т. К Megalonychidae принадлежал одним из наиболее широко распространённых видов — Megalonyx jeffersonii. Он достигал длины 3 м при весе более 1 т. Имеются свидетельства об охоте на M. jeffersonii человека (Research reveals ..., 2012). Megalonychidae вымерли около 5000 лет назад (Steadman et al., 2005). Ареалы наземных ленивцев были обширными. M. jeffersonii обитал от юга от Скалистых гор (найден на высоте 2700 м до Юкона на севере. Остатки E.laurillardi найдены от Южной Каролины до Бразилии.
Из непарнокопытных здесь жили настоящие лошади Equus три вида тапиров: калифорнийские (Tapirus californicus и T. merriami) и флоридский T. veroensis.
Разнообразны были и парнокопытные: два рода эндемичных близких к жирафам вилорогов Antilocapridae (один выжил), два рода оленей, в том числе оленелось Скотта Cervalces scotti, достигавший 2,5 м высоты и веса 710 кг, овцебыки, заселившие Северную Америку в раннем плейстоцене, задолго до Bison и давшие адаптивную радиацию. Кроме уцелевшего арктического вида (охота на него велась в материковой тундре Канады до 1917 г.) тут обитали Bootherium bombifrons (от Аляски до Калифорнии, вес более 400 кг) и Euceratherium collinum (от северной Калифорнии до центральной Мексики, вес до 600 кг). Полностью исчезли мозоленогие (Camelops hesternus, Hemiauchenia spp. иPalaeolama).
В состав голоценовых СС входили два вида бизонов Bison antiquus и B. latifrons.
Сайга Saiga tatarica (то же вид, что ныне обитает в евразийских степях) обитала в Северо-западных территориях Канады и на Аляске до 10 000 лет назад (Harington, 1998).
Гигантские пекари. Два рода (Platygonus и Mylohyus) небольших свиней, близкие родственники современных пекари. Отличались более крупными размерами (длиной в метр и более) и длинными ногами, благодаря которым животные были хорошими бегунами. Достигали веса более 150 кг. Более мелкий (до 90 см ростом, до 70 кг весом), наоборот, имел длинную вытянутую лицевую часть головы. Жили длинноногие пекари как в Южной, так и в Северной Америке, вероятно передвигаясь по прериям и редколесьям небольшими группами, наподобие антилоп. На север доходили до Юкона. Исчезли около 12—9 тысяч лет назад.
В состав мегафауны входили даже грызуны, например капибара Neochoerus pinckneyi весом до 114 кг и гигантские бобры Castoroides (широко распространённый C. ohioensis и эндем Флориды C. leiseyorum). Скелет в экспозиции Chicago's Field Museum имеет почти 2,5 м длины. Животное такого размера могло весить до 200 кг. Современный бобр не длиннее 1 м и весит до 30 кг. Резцы гигантских бобров были больше клыков саблезубых тигров. Гигантские бобры процветали от Флориды до Юкона и от Нью-Йорка до Небраски, но особенно в районе Великих озёр. Вымерли около 10 000 лет назад (Harington, 1996). Нет никаких свидетельств того, что гигантские бобры строили плотины, вероятно они этого не делали, поскольку плотины такого размера вряд ли остались незамеченными.
Как и в других районах Земли (за исключением очень холодных) в состав биоты Северной Америки входили крупные черепахи, принадлежавшие к Hesperotestudo. Карапакс H. crassiscutata и H. crassiscutataдостигал 1,25 м в длину (Rhodin et al., 2015).
Таким образом, биота Северной Америки была, в общем, сходна с биотой Евразии, в высоких широтах было много общих видов. Общая картина СС также была сходна: господствовали мозаичные ландшафты, в которых травоядные животные открытых пространств и олени, лошади (возникшие, кстати в Северной Америке), овцебыки, верблюды и ламы сосуществовали с гигантскими бобрами и другими обитателями леса на востоке США (Guilday, 1967, 1984 и др.). Некоторое отличие состояло в том, что в Северной Америке было гораздо выше разнообразие нежвачных фитофагов, жвачные же открытых пространств умеренных широт принадлежали к бизонам и вилорогам (другие жвачные жили в лесах).
С исчезновением кормовой базы резко сократили ареал и вымерли примерно 10 тысяч лет назад питавшиеся ими хищники. Лев Panthera leo (тот же вид, что ныне живёт в Азии и Африке), флоридский пещерный медведь Tremarctos floridanus, саблезубые кошки Smilodon fatalis и Homotherium serum (немного мельче Smilodon, до 1 м высотой, обитал преимущественно на севере Северной Америки, но заходил и в Южную. Вымер 10 тысяч лет назад), американский “гепард” Miracinonyx (2 вида). Другой крупный хищник — короткомордый медведь Arctodus simus достигал примерно 1,5—1,7 м в холке и веса до 1 т. Когда он становился на задние лапы, его высота достигала примерно 3,4 м (выше баскетбольной корзины!). Из аборигенных медведей Америки A. simus был самым молодым (поздним) и почти самым специализированным на хищничестве видом медведей (более хищный медведь только белый, но он живёт там, где другая пища малодоступна), его длинные ноги, вероятно, позволяли ему совершать длительные погони за добычей в прериях. Наиболее поздние находки A. simus в Америке датируют 10—11,5 тысяч лет назад. На юге Северной Америки A. simus сменял близкий вид A. pristinus. Об относительной числености крупных хищников можно судить по данным из битумных ловушек Ла Бреа (на территории США). На две тысячи смилодонов приходилось три тысячи Canis dirus (тот же вид, что и в Южной Америке) и лишь около 70 арктодусов. Вымерли и многие другие крупные хищники, включая, например, обычную пуму, ареал которой в Северной Америке восстановился совсем недавно. Вымерли также такие питавшиеся трупами крупных животных падальщики как кондоры (Gymnogyps amplus, Breagyps clarki) и их более крупный родственник Teratornis merriami.
Всего в Северной Америке в плейстоцене (после ледникового периода) вымерло 35 родов (55 видов) или 73% видов крупных млекопитающих (весом более 50 кг) (Edmeades, 2005). До сих пор сохранились деревья, для прорастания семян которых необходимо, чтобы они прошли по кишечнику. Однако не сохранились животные, столь большие, чтобы проглотить плоды этих деревьев.
Скорость преобразований биоты и их масштабы в Северной Америке в голоцене были гораздо больше, чем в северной Евразии и тем более в Палеотрописе. Исчезновение мегафауны произошло столь быстро, что в палеонтологической летописи не остались, да и вряд ли могли остаться свидетельства, как и почему это произошло. Синхронно с исчезновением мегафауны в Северной Америке появился человек, уже в совершенстве освоивший методы охоты на мегафауну; тогда как животные были незнакомы с человеком и не боялись его. В результате, если в африканской части Палеотрописа степень воздействия человека на биоту возрастала плавно на протяжении 2—3 миллионов лет, в азиатской и в северной Евразии — десятков тысяч, то в Северной Америке травоядные гиганты были уничтожены практически мгновенно, в основном всего за тысячу лет (см. выше).
Районирование Голарктики
Зональность. На Рис. 3.63—3.65 приведено несколько схем современной зональности Голарктики. Общая смена эдификаторов лесных климаксов с юга на север вечнозелёные дубы Quercus illes (подзона жестколистных лесов субтропической зоны) → листопадные дубы Quercus (зона летнезелёных лесов) → пихта Abies (подзона южной тайги) → ель Picea (подзона северной тайги), до того, как влияние человека стало заметным, вероятно, была той же, что и ныне. При рассмотрении распределения СС надо вспомнить последовательность сукцессионных стадий (о них даёт представление схемы сукцессий на Рис. 1.38 и 2.27) и обсуждение выше при рассмотрении тайги и широколиственных лесов биологической сути тундры и степей. Необходимо иметь в виду, что разделить подзоны тайги по имеющимся палеонтологическим данным невозможно, а субтропическая зона в Европе и ныне занимает узкую прибрежную полосу, в плейстоцене же её площадь была ещё меньше, и поэтому данных по этой зоне практически нет. Для северных районов в плейстоцене указывают сосновоберёзовые леса. Это, конечно, не следует понимать, как свидетельство существования смешанного леса из сосны и берёзы, поскольку сосна и берёза входят в разные сукцессионные ряды (ксеросерию и мезосерию), а лишь как указание на то, что эти виды — наиболее массовые в исследованных образцах пыльцы. Эти СС должны были заканчиваться зональным климаксом (ель, пихта или дуб) вне зависимости от того, найдены пыльца этих видов или нет, особенно это относится к дубу, который расселяется весьма медленно и сукцессии в СС с дубом в климаксе заканчиваются диаспорическим субклимаксом даже ныне (Главу 2). Представление о фауне млекопитающих Европы даёт Рис. 3.66. Формирование современной зональности Европы, преимущественно Восточной, происходило примерно так (Смирнова и др., 2004 а,б; Смирнова, 2006; Турубанова, 2002; Эволюция экосистем Европы..., 2008). Динамика ареалов эдификаторов различных зон и отдельных видов мегафауны отражена на Рис. 3.67-3.69.
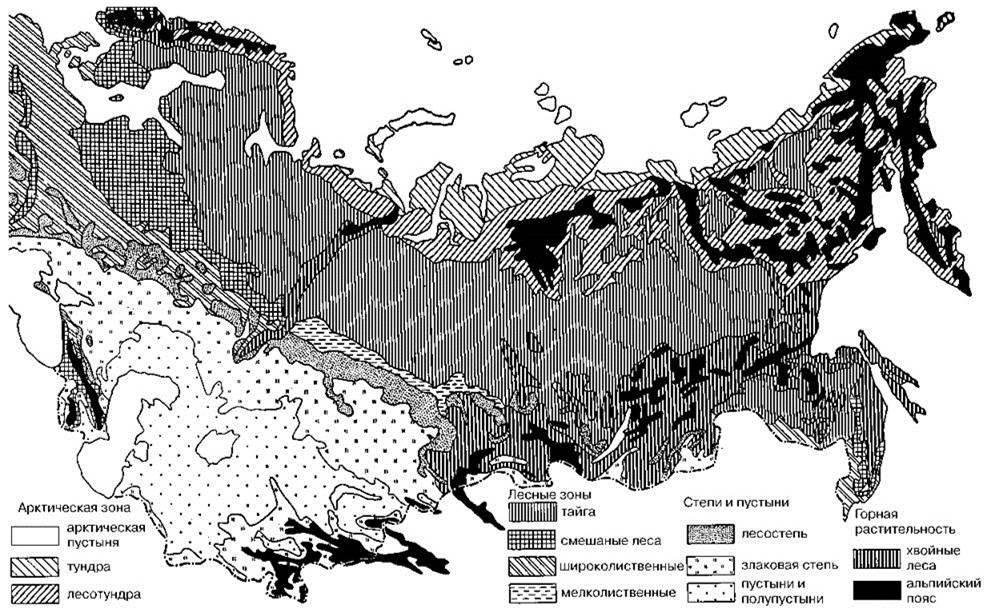
Рис. 3.63. Зональность растительности Евросибири по Г. Вальтеру (19756).
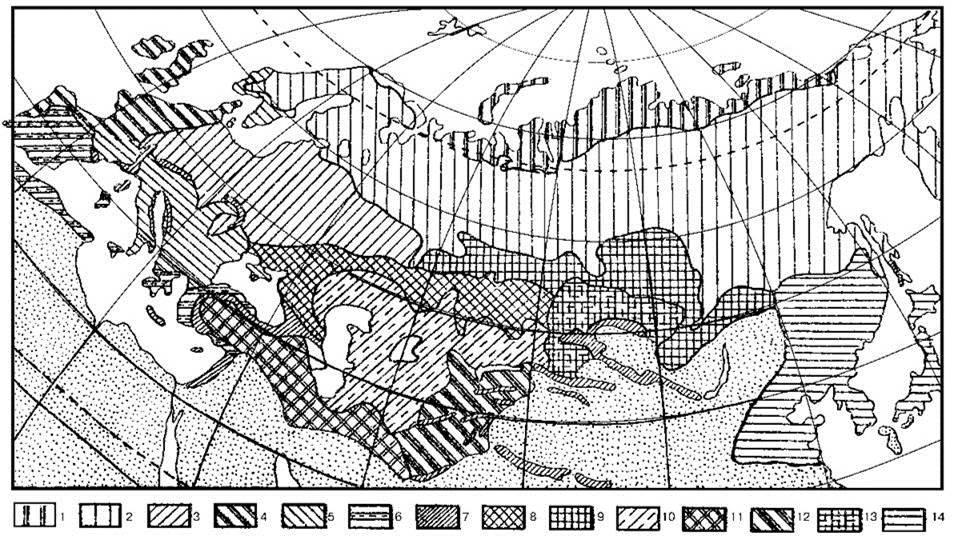
Рис. 3.64. Флористическая зональность Евросибири по Ю.Д. Клеопову (1990).
Регионы: 1 — Арктический; 2 — Бореальный; 3 — Среднеевропейский (в широком смысле); 4 — Атлантический; 5 — Субсредиземноморский; 5 — Средиземноморский; 7— Эвксикский; 8 — Понтический; 9 — Южносибирский; 10 — Туранский; 11 — Иранский; 12 — Центральносибирский; 13 — Северомонгольский; 14 — Восточноазиатский (включающий высокогорные хребты).
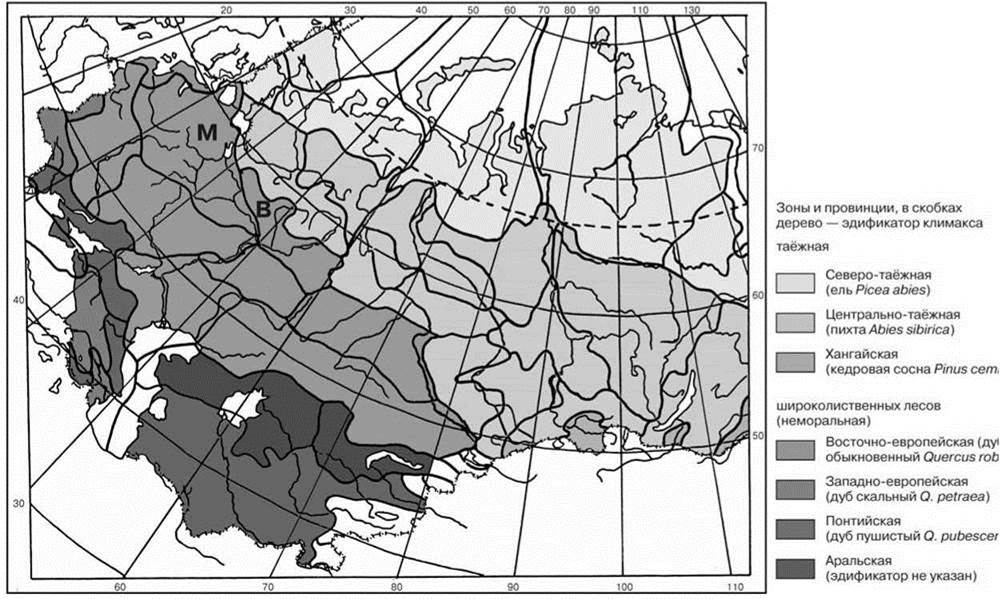
Рис. 3.65. Зональность северной Евразии по климаксным ассоциациям по С.М. Разумовскому (1999).
Сукцессионные схемы Московского (М) и Ветлужского (В) районов даны на рис. 1.38, 2.27

Рис. 3.66. Позднечетвертичная фауна млекопитающих Европы. По: Тенуис (1962) из Парфёновой (1999).
1 — лось Alces alces, 2 — гигантский олень Megaceros giganteus, 3 —выдра Lutra lutra, 4 — мамонт Mammuthus primigenius, 5 — лань Cervus dama, 6 — пещерный медведь Ursus spelaeus, 7 — северный олень Rangifer taran- dus, 8 — европейская косуля Capreolus capreolus, 9 — благородный олень Cervus elaphus, 10 — барсук Meles meles, 11 — леопард Panthera pardus, 12 — песец Alopes lagopus, 13 — бизон Bison priscus, 14 — тур Bos primigenius, 15 — европейский плейстоценовый осёл Asinus hidruntinus, 16 — росомаха Gula gulo, 17 — волк Canis sp., 18 — бурый медведь Ursus arctos, 19 — горный козёл Capra ibex, 20 — красный волк Cuon alpinus europaeus, 21 — овцебык Ovibos moschatus, 22 — шерстистый носорог Colodonta antiquitatus, 23 — серна Rupicapra rupicapra, 24 — пещерный лев Panthera spelaea, 25 — сайга Saiga tatarica, 26 — пещерная гиена Crocuta spelaea, 27 — лошадь Пржевальского Equus przewalskii, 28 — волк Canis lupus, 29 — кабан Sus scrofa, 30 — рысь Lynx lynx, 31 — лесной кот Felis silvestris, 32 — лиса Vulpes vulpes.
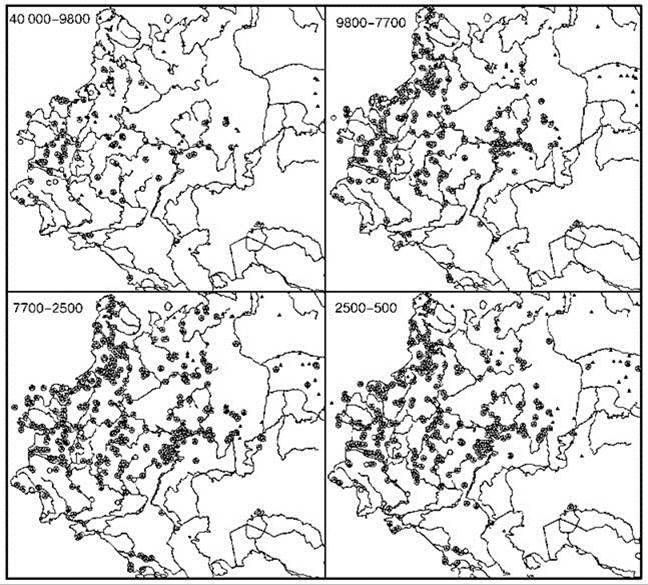
Рис. 3.67. Ареалы широколиственных (круги) и тёмнохвойных (треугольники) видов деревьев.
Широколиственные: дуб Quercus robur, бук Fagus sylvatica, клён Acer platanoides, ясень Fraxinus excelsior, липа Tilia cordata, вяз Ulmus minor, граб Carpinus betulus, лещина Corylusavellana. Тёмнохвойные: ель Picea abies и пихта Abies spp. По: Смирнова, Турубанова (2004).
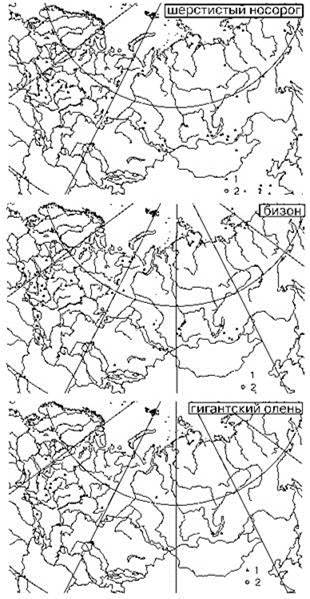
Рис. 3.68. Динамика ареала некоторых видов макрофауны.
Находки: 1 — позднего плейстоцена; 2 — голоцена. По Калякину, Турубановой (2002).
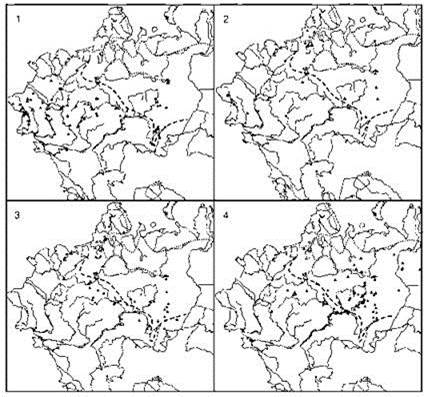
Рис. 3.69. Динамика ареала северного оленя.
Находки: 1 — поздний плейстоцен — древний голоцен; 2 — ранний голоцен; 3 — средний голоцен; 4 — поздний голоцен по Калякину, Турубановой (2002). Границы ареала по Гептнеру и др. (1961): пунктир — восстановленная в историческое время, точки — середина 1950-х гг.
Максимальная стадия последнего оледенения. Внутри этого интервала, между 22-18 тысяч лет, Скандинавский ледниковый щит существенно увеличился и достиг максимальных для позднего плейстоцена размеров. В результате роста оледенения ко времени его максимального развития уровень моря понизился на 120 м относительно современного (Bard et al, 1990; Sidall et al., 2003). По побережьям у ледника в этот период и позднее встречался белый медведь. В узкой полосе, прилегающей к ледниковому щиту — лесотундре — господствовали ранние гидросериальные стадии тайги с мозаикой березовых и сосновых лесов. Южнее лесные стадии тайги были более обычны, Пыльца доминантов климакса тайги (ели и пихты) встречается практически от края ледника, но, как и следовало ожидать, у ледника в небольших количествах. Большая часть видов мамонтового комплекса (шерстистый носорог, гигантский олень, сайга, первобытный бык, первобытный бизон, овцебык, гиена, лев, пещерные медведи) в лесотундре отсутствовала, на юг они были распространены до примерно широты северных берегов Чёрного моря. Южнее этой границы влияние ледника было практически не ощутимо, и господствовали неморальные СС. Впрочем, отдельные виды, характерные для неморальных лесов, шли на север почти до современных границ их распространения; предполагают, что они сохранялись на территории рефугиумов. Повсеместно распространены типичные степные и даже некоторые пустынные виды растительности, а также соответствующие виды животных (сайга, лошади, бизоны). Это служит дополнительным подтверждением отсутствия сплошных лесных зон. В это время в Европе вымирают большой и малый пещерные медведи и пещерная гиена. Более южные виды (слоны и бегемоты) к этому времени на территории континентальной Европы уже исчезли. Последние неандертальцы вымерли до 25 тысяч лет назад (в Южной Испании и, возможно, в Хорватии (Верпоорте, 2008).
Позднеледниковье (<17,0 — >=12,4 тысяч лет назад). Распределение СС в общих чертах повторяет предыдущий период, но в результате хода сукцессий в каждой зоне увеличилась роль следующих сукцессионных стадий.
Потепление бёллинг-аллерёд (<=12,8 — >10,9 тысяч лет назад). К началу этого временного интервала Скандинавский ледниковый щит сильно деградировал, но все еще существовал (Lundqvist, 1986). Его восточная часть, на северо-западе России, быстро таяла и отступала со средней скоростью около 200 м в год. В конце периода ото льда освободились такие крупные озера как Онежское и Ладожское и южная часть Белого моря (Saarnisto, Saarinen, 2001). Для этого времени характерно интенсивное торфонакопление и развитие почвенного покрова на значительной части Европы. Сплошных лесных поясов не было, неморальная растительность была распространена значительно южнее современной границы. Виды мамонтовой мегафауны становятся более редкими и границы их ареала сдвигаются на север и/или восток.
Поздний дриас (<=10,9 — >10,0 тысяч лет назад). Короткий период похолодания. Вновь возникли ледники в Скандинавии и Шотландии. Сплошные пояса лесов отсутствуют. Виды мамонтового комплекса сохранились в виде реликтовых популяций в наименее населённых районах.
Пребореал и бореал = ранний голоцен — мезолит (<=10,0 — >=8 тысяч лет назад). Ледники начали таять и около 8500 лет назад исчезли окончательно (Lundqvist, 1986). Разрозненные реликтовые популяции видов мамонтового комплекса исчезают. Вероятно, в этот период вымирают мамонт и многие кавказские виды — кавказский сурок, медведь Денингера, красный волк; очень возможно, что вымирают овцебык, шерстистый носорог, бизон; пещерный лев. Еще позднее вымирает гигантский олень. Роль эдификаторов переходит к крупным копытным (зубр, тур, лось, торфяной олень, разные виды лошадей). Эти виды подавляли развитие лесной растительности, хотя и не столь эффективно, как хоботные и носороги, но всё же препятствовали развитию сплошных лесов путём создания более или менее обширных луговых и лугово-степных зоогенных полян. Комплексы млекопитающих, характерные для современной тайги, широколиственных лесов и лесостепи, еще не сформировались Характерным видом тайги являлся песец, позднее, в связи с уменьшением площадей, занятых ранними гидросериальными стадиями тайги (“тундрой”), сохранившийся только на севере таёжной зоны — “тундре”. Наиболее поздние находки мамонта в Европе сделаны на севере Русской равнины и в Прибалтике, что свидетельствует о том, что этот вид входил в состав неморальной териофауны. Виды млекопитающих, которых ныне считают степными обитали и в наиболее северных из изученных районов (как минимум до Карелии): это свидетельствует об отсутствии сплошных зон лесов и в этот период. В комплекс млекопитающих, характерный для южных районов, ныне занятых степями, наряду с сайгой, лошадьми, бизонами и быками входили благородные олени, кабаны и бобры, что свидетельствует о значительной облесенности степей. Конец мезолита примерно совпадает с завершением ландшафтной перестройки, после чего для обеспечения всё возрастающих потребностей увеличивающегося населения произошла следующая техническая революция — было изобретено земледелие (Ламберт, 1991).
“Основные изменения в составе фауны млекопитающих Европы произошли в голоцене, а не в конце плейстоцена” (Косинцев, 2008: 268).
“Большинство таёжных ландшафтов относительно слабо нарушено хозяйственным воздействием” (Исаченко, Шляпников, 1989: 63).
В среднем голоцене (7,7—2,5 тысяч лет назад). Хозяйственная деятельность человека, связана уже не только с охотой, но и с сельским хозяйством, что позволило резко возрасти общей численности населения. Из-за увеличения охотничьих усилий численность крупных стадных копытных и бобров была снижена настолько, что они потеряли свою эдификаторную роль, поймы рек приобрели современный облик, а бобровые пруды исчезли. Неизбежно возросшая сухость климата Восточной Европы привела к ухудшению произрастания темнохвойных деревьев и сдвиганию южной границы их ареала на север. Выжигание и вырубание лесов на севере привело к расширению тундры, а в более южных районах — к широкому распространению сосновых боров и безлесных районов. Как известно, подсечно-огневая система земледелия наиболее древняя. При её использовании землю не надо было обрабатывать, т. е. не нужны ни какие-либо орудия, ни тягловая сила, поэтому, по мнению археологов, такое земледелие возникло задолго до появления железного топора. Деревья не срубали (поскольку было нечем), а окоряли, чтобы они засохли. Подготовленный участок выжигали таким образом, чтобы лес, сгорая, выжег почву на глубину как минимум 5 см, полностью уничтожая всю растительность, в том числе и семена. Посев производили в ещё тёплую золу, в этом случае на поле сорняки возникали только из засорения посевного материала. Выбор участков производили по растительности, наиболее ценными были земли, занятые климаксными лесами — дубравами (Советов, 1867; Петров, 1968). Участки забрасывали не из-за истощения почвы, а из-за зарастания полей сорняками, с которыми могли бороться только прополкой — способом неэффективным на больших площадях, ибо урожай не окупал затрачиваемый труд. Большие затраты труда на расчистку окупали высокие урожаи: до сам-10 — сам-20, а иногда и до сам- 70 (!) (Милов, 1998). Площадь климаксных лесов в северных районах была незначительной, поэтому их быстро вырубили. Масштабы подсечно-огневого земледелия не следует недооценивать: даже в такой малонаселенной стране как Финляндия (зона тайги), за XVIII—XIX вв. через подсеку прошло 85% территории (Смирнова и др., 2001). Расселение дуба, лещины и других широколиственных деревьев происходит медленно, поэтому диаспорические субклимаксы становились всё более обычными, и северная граница неморальных лесов постепенно сдвигалась на юг. Полное исчезновение на севере видов, разрушающих моховой покров, привело к его разрастанию и формированию пояса тундры. Проводимые Зимовым опыты показывают, что разрушение мохового покрова копытными (лошадьми) приводит к замене мохового покрова злаковой степью и резкому росту продуктивности растительности и всей экосистемы. Широкое распространение на юге подсечно-огневого земледелия и скотоводства привело к уничтожению лесов и формированию на их месте степной и полупустынной зон. Уничтожение лесов происходило не только в результате вырубки лесов, но и из-за выпаса скота, уничтожавшего подрост, а также использования лесной дернины для подстилки в стойлах скота (отсюда, вероятно, и двоякое значение слова “подстилка”: словарь Брокгауза и Ефрона различает лесную и хлевную подстилки). В результате начинается антропогенное расслоение единого ранее пояса на четыре: тундру, бореальные (тайга) и неморальные (широколиственные) леса и степь, к которым позднее добавили кукурузный и другие пояса.
Ранний железный век и средневековье (500—2500 л.н.). В начале позднего голоцена — в раннем железном веке (1600—2500 л.н.) ареалы древесных видов ещё не претерпели существенных изменений, широколиственные и темнохвойные виды практически всюду встречаются вместе. Но по сравнению со средним голоценом доля пыльцы темнохвойных видов сокращается на юге и увеличивается на севере (Нейш- тадт, 1957). Это свидетельствуют о начале трансформации ареалов темнохвойных и широколиственных видов (Нейштадт, 1957; Хотинский, 1977; Восточноевропейские..., 1994). Массовые рубки для выплавки железа и подсечно-огневое земледелие приводили к значительному уменьшению лесистости обширных районов и было мощным фактором аридизации, особенно на юге, и привели к формированию пояса степей. Значительные перемещения на север южной границы ареалов ели и пихты в условиях более холодного и влажного климата позднего голоцена (по сравнению со средним голоценом), видимо, можно объяснить только антропогенной аридизацией юга Восточной Европы. Резко возросшие потребности металлургии и судостроения привели к перемещению на юг северной границы ареала дуба (Гмелин, 1771; Паллас, 1773).
Судя по находкам крупных деревьев или их пней леса росли на берегах Северного Ледовитого океана ещё в XVIII—XIX вв. (Городков, 1954), а на Ямале — в XIX в. (Калякин, 1985, 1991). Оленеводы регулярно устраивали палы для сведения лесов вплоть до XVIII в. (Шренк, 1855). В результате к XVUI—XIX вв. сформировался пояс тундры.
Фауна большей части Восточной Европы имела смешанный характер, местами вплоть до недавнего времени. Так, в конце XVIII в. на пространстве от нынешней Воронежской области до севера Казахстана и юга Западной Сибири еще встречались тарпаны и куланы (Гмелин, 1771); северные олени при зимних кочевках регулярно доходили до севера калмыцких степей (Паллас, 1773) и до Украины (Гепнер и др., 1961). В начале XIX в. сурки были распространены до нынешней Калужской области (Зельницкий, 1804), а в конце XVIII в. были обычны в Тамбовской губернии (Паллас, 1773).
На островах Средиземного моря (Корсика, Сардиния, Сицилия, Балеарские острова, Мальта, Крит, Кипр), климат которых вряд ли существенно отличался от климата материковых побережий Средиземного моря, но на которых не было людей, плейстоценовая биота сохранилась гораздо дольше, хотя и в модифицированном из- за малых размеров островов виде. Виды мегафауны дали карликовые формы на каждом из островов, часто выделяемых в отдельные виды. Выше уже было сказано о карликовых слониках. Бегемот (вид, идентичный современному или очень близкий) дал островных бегемотиков размером со свинью. Аналогичные карликовые формы дали и торфяные олени (<1 м и даже 0,6 м в плечах и очень маленькими рогами) и быки. При этом уменьшение размеров сопровождалось непропорционально сильным укорочением ног, поскольку из-за отсутствия крупных хищников необходимость в быстрых передвижениях отпала (Agusti, Anton, 2002). После того, как острова были открыты человеком и леса были вырублены полностью, вся эта фауна исчезла.
В других районах Голарктики аналогичные изменения могли происходить в иное время и несколько иначе. В частности, Сибирь была зоной процветания типичной “мамонтовой фауны” и была богаче видами, чем сегодняшний и даже прошлый гористый и южный Дальний Восток (где в голоцене выпало не так много животных, только мамонт и бизон, да пещерная гиена). Выше была и численность плейстоценовой макрофауны. После разрушения мамонтовых СС в конце плейстоцена и в Сибири начала формироваться отчётливая широтная зональность с зарастанием и заболачиванием прежних мозаичных ландшафтов. Возникла огромная территория бедной тайги шириной в сотни километров и длиной в тысячи и тысячи. СС представлены там преимущественно ассоциациями пирогенных рядов с лиственницей и сосной. Широкое распространение пожаров, из года в год горящих в Сибири, связано как со сменой “клочковатого” ландшафта однородными лесами, так и уничтожением мегафауны, потреблявшей ресурсы, ныне питающие преимущественно огонь. От прежнего разнообразия из крупных животных остались бурые медведи, мелкие лоси (вид исходно лесостепной), северные олени и волки, местами рысь... Севернее произошло формирование обширного болота на вечной мерзлоте (тундры).
Биогеографические районы. Несостоятельность разделения Голарктики на Палеарктику и Неарктику с биологического точки зрения обсуждена выше.
Биота Голарктики образовалась в результате процессов, аналогичных тем, которые привели к образованию биогеографических районов на юге Южной Америки и Африки: адаптации биоты Евразии и Северной Америки, соединённых Беринговым перешейком в единый континент ко всё более холодному климату. Как и в остальных случаях, в биоту умеренных районов не смогли войти многие таксоны, поэтому биоту Голарктики характеризуют не столько её эндемы, сколько отсутствие широко распространённых в тропиках таксонов (необходимо помнить, что многие из них, в первую очередь мегафауна — слоны, носороги, бегемоты, крупные кошки, страусы и др. — истреблены в Голарктике человеком). Число эндемов Голарктики, распространённых как в Евразии, так и в Северной Америке, невелико. Из млекопитающих это кроты Talpidae, бобры, прыгунчики Zapodidae, пищухи, или сеноставки Ochonotidae. В мегафауне (как вымершей и истребленной, так и сохранившейся) эндемизм не превышает родового уровня. Из птиц: тетеревиные, гагары и чистиковые, впрочем, последние два семейства логичнее рассматривать как компоненты морской, нежели сухопутной биот. Существенную роль в почвах Голарктики играют эндемичные дождевые черви Lumbricidae. Эндемичны сосновые Pinacea.
Конфигурация фрагментов (от одного до трёх в разные периоды времени) суши, на которых расположена Голарктика, существенно изменялась (описание событий дано в следующей Главе). Дольше всего единство суши, обеспечивавшей обмен биотами между разными её частями, сохранялся на севере, тогда как южные районы развивались в значительной степени в изоляции. Это сделало возможным образование большого числа таксонов в их южный частях. Таксоны мегафауны, возникшие в Северной Америке (лошади Equidae и верблюды Camelus************), проникли в Евразию и даже в Африку. Таксоны, включающие более мелкие по размерам виды остались эндемами или субэндемами (проникнув в Южную Америку): вилорогие антилопы Antilocapridae*************, бобровые белки Aplodontidae, мешетчатые крысы, или гоферы Geomyidae, мешетчатые мыши Heteromyidae, еноты Procyonidae. Большое число эндемичных таксонов, возникших в Северной Америке, объясняется тем, что евразийская часть Голарктики всё время находилась в непосредственном контакте с гораздо более таксономически разнообразной биотой Палеотрописа. Это не способствовало образованию эндемов (среди млекопитающих эндемичны лишь слепыши Spalacidae и тушканчики Dipodidae). Вместо этого происходило обогащение биоты палеотропическими таксонами, часть из которых (парнокопытные, хоботные, кошачьи, собачьи, медведи) смогла проникнуть и в Северную Америку. Сохранявшаяся почти непрерывно сухопутная связь северных районов Евразии и Северной Америки то через Гренландию, то через Берингов перешеек привела к образованию весьма однородной биоты северных районов Голарктики, тем более однородной, чем с более северными районами мы имеем дело. К тому же в районах, покрытых во время последнего оледенения ледником (север Европы, вся Канада) жизнь восстановилась лишь в последние 10 000 лет или чуть более. Аляска была населена преимущественно азиатской биотой, поскольку она была соединена с ней широким сухопутным мостом и отделена ледником от остальной части Северной Америки, впрочем некоторые северо- и даже южноамериканские таксоны сохранились там с предшествующих межледниковий. В результате на севере зоны тайги биоты Северной Америки и Азии неразличимы, на юге зоны тайги сходство велико, но есть и специфика, а, начиная с зоны широколиственных лесов, различия превышают сходство. Но и на севере зоны широколиственных лесов многие рода общие, представленные разными видами. И лишь южнее различия биот обусловлены таксонами ранга рода и семейства и даже более высокого. Различие биот Северной Америки и Азии усилилось после образования Панамского перешейка. Его возникновение позволило многочисленным таксонам Неогеи проникнуть на север (обычно до зоны тайги), но не в Азию. Это даёт основания разделить Голарктику на четыре подобласти: Бореальную, Средиземноморскую, Восточноазиатскую и Сонорскую. Последнюю, возможно, следует разделить на восточную и западную надпровинции, соответственно восточно- и западноамериканским флорам (см. Главу 1). Это деление грубо соответствует современным ареалам зоны тайги, и трёх изолированных фрагментов зоны широколиственных и субтропических лесов. Соответствие это не полное, поскольку современные границы этих зон в значительной степени — результат деятельности человека. Было ли это совпадение до изменения облика суши человеком более полным или нет, сказать в настоящее время практически невозможно. Давать более детальное районирование Голарктики по этой же причине вряд ли целесообразно.
* Разные авторы использовали разные названия рангов выделенных ими фаунистических регионов. Для облегчения сравнения схем, независимо от названий рангов, данных выделам авторами схем, в этой и остальных главах самые крупные выделы названы областями, более мелкие — подобластями и провинциями.
** Первые исследователи полагали, что этими клыками свинья цепляется за ветки, давая голове отдохнуть, а у самок их нет, поскольку потребность самок в отдыхе не столь велика.
*** Среди австралийских сумчатых нет и водных видов, единственное водное млекопитающее Австралии — утконос, в Новой Гвинее воду освоили опять-таки Muridae. Впрочем в Южной Америке водные сумчатые есть.
**** Ныне всего один вид, второй вымер в 1965 г.
***** Создаётся впечатление, что в островных СС роль верховного хищника, как правило, переходит к гигантским орлам. Вымершие в конце плейстоцена крупные хищные птицы были описаны из Вест-Индии и даже со средиземноморских островов (Naish, 2008), недавно был описан Stephanoaetus mahery Goodman, 1994 с Мадагаскара, сходный с орлом Хааста. С появлением человека, ситуация, естественно, резко менялась.
****** Формирующийся человек в Африке застал 44 вида животных весом более 1 т (Barkham, 2016).
******* Данная публикация имеет некоторые неточности. Так, авторы ошибочно отнесли медведей Arctotherium ко второй группе, хотя их вес существенно превышал 1 т, а Canis dims — вид заметно более крупный, чем современный волк C. lupus, к животным весом <44 кг, хотя вес C. lupus может достигать почти 90 кг. Также они считают, что в Южной Америке обитало 5 видов хоботных, тогда как специалисты по этой группе (Prado et al., 2001) признают валидность только трёх.
******** Два вида дожило до прихода человека: (1) S. fatalis (высота до 1,2 м, вес до 300 кг, верхние клыки достигали длины 18 сантиметров), до прихода человека — самый массовый и широко распространённый крупный хищник Южной и Северной Америк, и (2) S. populator, обитавший на востоке Южной Америки (крупнейший вид Felidae, достигал 1,4 м в плечах, 2,6 м длиной с 30 см хвостом, вес, вероятно, превышал полтонны).
********* Отдельный подотряд хоботных, противопоставляемый остальным (мастодонтам, слонам, мамонтам, стегодонам и др.). Отличались отсутствием бивней в верхней челюсти и наличием изогнутых вниз и назад бивней в нижней. Возникли в Африке, обитали и в Западной Европе. Никогда не были разнообразны (Shoshani, Tassy, 2005). Наиболее поздние находки — 1 млн. лет назад (Кения).
**********Grubb et al. (2000) показали морфологическую разницу между двумя подвидами Loxodonta africana, достаточную, по их мнению для повышения ранга подвидов до видового. Roca et al. (2001) показал, что молекулярные данные подтверждают это мнение. Однако Eggert et al. (2002), также основываясь на молекулярных данных показали, что нет различий между L. cyclotis и L. africana, а есть различия между географическими группами, не соответствующими группам, выделяемым по морфологии.
*********** По другим данным на Мадагаскаре эпиорнисы сохранялись до XVIII—XIX вв. Ископаемые останки эпиорнисов известны и из неогеновых отложений Передней Азии и Северной Африки. Вероятно, эпиорнисы и гигантские орлы стали прообразами гигантской птицы Рух арабских сказок “Тысячи и одной ночи”. Имеются устные сообщения местных жителей о том, что ещё в 1976 г. в одной из отдалённых деревень жил одомашненный бегемот.
************ В Южную Америку проникли только в плейстоцене (Дарлингтон, 1966). В Евразию верблюды рода Paracamelus проникли в нижнем плиоцене, или немного раньше.
************* Известны из нижнего плиоцена Монголии (Дарлингтон, 1966).