Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Озеро Танганьика - Специфика биот крупных озёр - Общая характеристика континентальных водоёмов как биотопа - Биогеография континентальных водоёмов
Ныне — озеро бассейна р. Конго, связь с бассейном р. Конго устанавливалась неоднократно, ранее вместе с оз. Ньяса входило в систему р. Замбези. Это самое старое и глубокое из великих африканских озёр. В глубинах озера кислород отсутствует; нижняя граница аэробной зоны расположена в среднем на 70 м на севере и на 240 м на юге. Термоклин расположен примерно на глубине 40—50 м. Температура на поверхности может превышать 27°С, на глубине 1 км — около 23°С (Coulter, Spigel, 1991). Впервые водоёмы на месте современного озера начали формироваться примерно 14 млн. лет назад, в миоцене. Это были мелкие маленькие изолированные озёра. Примерно 5 млн. лет назад (с середины плиоцена) рифт, в котором расположено озеро, начал интенсивно развиваться и озёрная котловина постепенно углублялась, что привело примерно 2 млн. лет назад к формированию глубоководного озера. В плейстоцене (последние 1,5 млн. лет) уровень озера временами существенно понижался, при понижениях до 400 и 600 м относительно современного уровня озеро распадалось на два. Но даже меньшие падения уровня озера приводили к существенному сокращению мелководий и, вероятно, частичному вымиранию озёрной биоты. Последнее существенное понижение уровня озера (на 300 м относительно современного уровня) было 16—14 тыс. лет назад (Coulter, 1991; Tiercelin, Mondeguer, 1991).
Из примерно 200 видов обитающих здесь рыб 95% — эндемы (Roberts, 1975). Наиболее высок эндемизм среди цихлид (вероятное число эндемичных видов цихлид оценивают в 325 (Leveque et al., 2008). У обитающих в Танганьике цихлид строение зубов выходит за пределы вариабельности даже не семейства, а всего класса (Рис. 4.6). Кроме того, здесь имеются эндемичные рода и виды семи других семейств рыб, особенно для Танганьики (но не для других великих африканских озёр) характерно разнообразие Bagridae, Mochokidae, Mastacembelidae и Centropomidae. Богатой эндемичной ихтиофауне соответствует богатая эндемичная фауна паразитов рыб. Из пяти родов Unionidae, обитающих в Африке, два — эндемы Танганьики (ещё один обитает в верхнем течении р. Конго, вытекающем из оз. Танганьика), имеются здесь эндемичные виды и широко распространённого Caelatura; эндемичны три вида Mutelidae. Фауна гастропод насчитывает 83 вида, из которых 65 — эндемичны, наиболее разнообразны Paludomidae,образовавшие 18 эдемичных родов (Strong et al., 2008). Эндемичны три рода креветок (Limnoca- ridina, Caridella и Atyella), 8 видов крабов. Эндемичных насекомых мало, в частности описано два эндемичных рода водяных клопов (Paskia и Idiocoris), но клопы озера изучены ещё недостаточно (Polhemus, Polhemus, 2008), все эндемичные насекомые, за исключением двух пелагических ручейников приурочены к прибрежью. Из 69 видов копепод 34 (49%) — эндемы (Coulter, 1991; Box-shall, Strong, 2006).
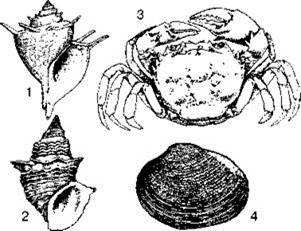
Рис. 4.6. Некоторые эндемы озера Танганьика
1 — Paramelania damoni, 2 — Tiphobia horei; 3 — Potamonautes (Platythelphusa) armata, 4 — наяда Caelatura burtoni (по Coulter, 1991).
Табл. 4.7. Эндемы некоторых таксонов в разных частях оз. Танганьика: число (процент от фауны данной части) (по: Coulter, 1991).
|
пелагиаль и глубоководная биота |
мелководная биота |
ассоциированные водоёмы (лагуны, марши, дельты рек и т.п.) |
||
|
губки |
2 (100%) |
7 (100%) |
2 |
(0%) |
|
мшанки |
3 (67%) |
4 (25%) |
0 |
|
|
Gastropoda |
13 (92%) |
35 (97%) |
25 |
(0%) |
|
Bivalvia |
0 |
8 (100%) |
7 |
(14%) |
|
Cladocera |
0 |
6 (0%) |
23 |
(0%) |
|
Copepoda |
||||
|
Calanoida |
1 (0%) |
0 |
0 |
|
|
Cyclopoida |
7 (43%) |
43 (33%) |
9 |
(33%) |
|
Harpacticoida |
0 |
16 (88%) |
0 |
|
|
Ostracoda |
0 |
56 (100%) |
31 |
(68%) |
|
Decapoda |
||||
|
креветки |
3 (100%) |
14 (100%) |
1 |
(0%) |
|
крабы |
5 (100%) |
7 (100%) |
2 |
(0%) |
|
рыбы |
||||
|
Cichlidae |
66 (100%) |
149 (100%) |
14 |
(64%) |
|
не Cichlidae |
21 (90%) |
59 (73%) |
72 |
(15%) |
|
Insecta* |
2 (100%) |
58 (16%) |
44 |
(14%) |
* В фауне озера отмечено 155 видов (12% фауны), но данные по распределению по анализируемым районам приведены только для 94 видов.
Приведённые в начале раздела цифры, как и в случае Байкала, характеризуют биоту озера в его географических границах. Анализ видовых ареалов показывает, что в пределах озера можно выделить те же три ЦС, что и в Байкале: мелководную (населяющую эпилимнион), глубоководную (населяющую гиполимнион) бентические и пелагическую. Плюс к тому здесь, как и в Байкале, прибрежные лагуны, заливы, марши и дельты впадающих в оз. Танганьика рек, географически относимые к озеру (associated water bodies), биогеографически и экологически к нему не принадлежат. Их населяет биота, мало отличающаяся от биоты окрестных водоёмов со сходными условиями (Табл. 4.7). Среди макрофитов озера нет ни одного эндема. Биота многих таксонов остальной части озера почти целиком эндемична, в других (например, Cladocera) эндемов нет нигде. Среди дышащих воздухом гастропод (прудовики Lymnaeidae, катушки Planorbidae, ампулярии Ampullariidae = Pilidae) также нет ни одного эндемичного вида, все они обитают в ассоциированных водоёмах. Эндемы имеются только среди дышащих жабрами гастропод, почти все они обитают за пределами ассоциированных водоёмов: из 36 эндемичных видов гастропод только четыре встречаются в ассоциированных водоёмах, и только три — исключительно в них. Согласно сводке Coulter (1991) из 4 подсемейств переднежаберных гастропод Thiaridae два эндемичны для Танганьики и оба они не встречаются в ассоциированных водоёмах. Поэтому мнение о происхождении эндемов на ранней стадии развития озера, когда на месте глубоководного озера существовало множество мелких изолированных озёр (см. Coulter, 1991) крайне малоправдоподобно: условия в таких озёрах соответствовали ассоциированным водоёмам, которые ныне населены широко распространённой в тропической Африке биотой, тогда как эндемы для них нехарактерны. Предполагать, что ранее всё было наоборот, нет никаких оснований. В то же время следует отметить, что некоторые таксоны моллюсков, ныне эндемичные для озера, ранее были распространены гораздо шире и найдены в отложениях соседних озёр. Размеры этих озёр с тех пор значительно уменьшились, что, вероятно, и послужило причиной обеднения их биоты.
Постоянная стратификация озера ведёт к медленному возврату биогенов из гиполимниона в эпилимнион, хотя во время сухого сезона на юге озера происходит подъём глубинных вод из-за сильных сгонных ветров (апвеллинг). Время полного обмена воды озера оценивают в 1000 лет, тогда как в Байкале вертикальное перемешивание происходит каждый год. Тем не менее, уровень первичной продукции фитопланктона в пелагиали озера примерно вдвое больше, чем в Байкале, около 300 гС/кв. м в год (Hecky, 1991), что позволяет характеризовать озеро Танганьику как эвтрофное. Необычно высока роль в фитопланктоне (по числу видов и биомассе) хризофитовых водорослей. В формировании первичной продукции озера, вероятно, существенную роль играют инфузории Strombidium cf. viride. Инфузории этого рода — фитофаги, питающиеся водорослями, но хлоропласты проглоченных водорослей Strombidium включают в свою протоплазму, где они долго функционируют. Strombidium озера Танганьика имеют в своих клетках водоросли, которые, возможно, являются их симбиотами. Биомасса Strombidium достигает 0,5 г/куб. м и даже выше, временами в несколько раз превышая биомассу всех водорослей вместе взятых. Особенно много Strombidium в дождливый период, когда стратификация максимальна (Hecky, 1991). В гетеротрофном зоопланктоне доминируют копеподы — широко распространённая Tropodiaptomus simplex (взрослые стадии — хищники) и несколько видов мелких Cyclopoida. Cladocera, как и в Байкале, полностью отсутствуют. Крупный зоопланктон образуют несколько видов эндемичного рода креветок Limnocaridina и гидромедуза (эндем или нет, в зависимости от таксономиста). Здесь нет коретры (Chaoborus), потребляющей существенную часть первичной продукции пелагиали других тропических озёр, в частности озера Ньяса. Продуктивность гетеротрофного зоопланктона оценивают в 50—60 г С/кв. м в год, что составляет около 20% продуктивности фитопланктона (Hecky, 1991). Пелагические рыбы Танганьики, в отличие от Байкала, не образовали отдельного эндемичного семейства, а лишь эндемичные рода. Вряд ли это стоит объяснять различием в возрасте озёр. Причина скорее в другом. В Байкале пелагическими стали исходно бентосные бычки, это потребовало их существенного изменения. В Танганьике же пелагические рыбы принадлежат к исходно пелагическим сельдям Clupeidae. Они относятся к двум родам, один из которых танганьикский “шпрот” Stolothrissa tanganicae эндемичен для озера, а другой — танганьикская “сардина” Limnothrissa miodon — встречается ещё только в верхнем течении р. Конго, гидрологически связанном с озером. S. tanganicae более мелкий вид, продолжительность жизни 99% особей не превышает одного года. S. tanganicae становится половозрелым при размере 75 мм (самки) и 64 мм (самцы). Молодь (менее 50 мм) питается фитопланктоном, более взрослые переходят на питание копеподами и креветками, день они проводят в гиполимнионе, ночью поднимаются на поверхность. L. miodon гораздо крупнее, максимальная его длина 175 мм, продолжительность жизни 2—3 года. Верхний трофический уровень пелагической ЦС образует два вида Lates(Centropomidae): L. microlepis и L. stappersi. L. microlepis достигает 80—100 см, L. stappersi — полуметра. В озере обитают ещё два вида этого рода, также эндемичные, но их основным объектом питания являются придонные рыбы (то, что они не быстрые пловцы, видно даже по их хвостовым плавникам, лишённым выемки). Впрочем, они способны подниматься по ночам в поверхностные слои для питания сельдёвыми. Протяжённость вертикальных суточных миграций может достигать 200 м. Продолжительность жизни всех видов Lates более 10 лет. Продуктивность рыб пелагиали озера оценивают в 650—700 кг/га в год (= 65—70 г/кв. м в год). Схема пищевой цепи пелагиали озера показана на Рис. 4.7.
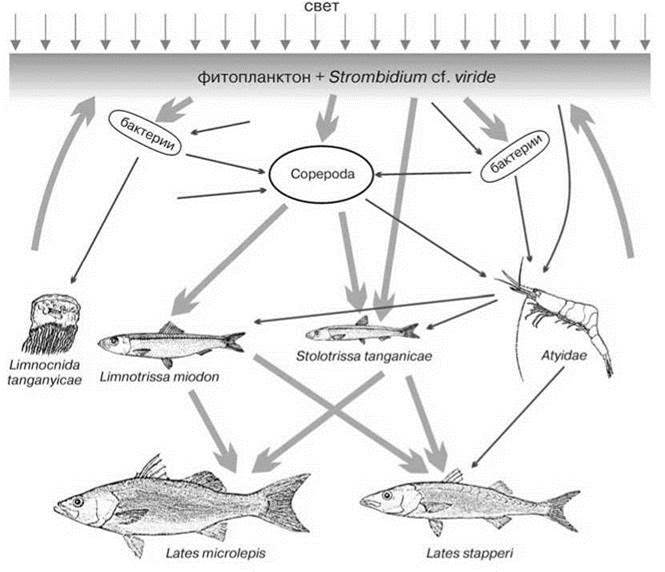
Рис. 4.7. Схема пищевых связей пелагиали озера Танганьика (ориг., по данным Heckey, 1991).
Толстые линии — главные связи, тонкие — второстепенные.
Оба вида сельдёвых являются основными промысловыми видами рыб озера. Limnothrissa miodon успешно акклиматизировали во многих африканских озёрах и водохранилищах, но там она не достигает столь крупных размеров, как в Танганьике. Промысловыми являются также все четыре эндемичных вида Lates. Общий объём промысла рыб в озере достиг в 1995 г. 180 тыс. т, и в нём занято более 100 000 человек, выходящих в озеро на более 12 тысячах каноэ, около 1000 крупных лодок и около сотни более крупных специализированных судов. Мелких сельдёвых после вылова сушат, раскладывая на пляже, сушёных рыб транспортируют на несколько тысяч километров.
В пелагической ЦС Танганьики, как и Байкала есть виды, связанные с поверхностной плёнкой (Рис. 4.3) — гипонейстон. Хотя карповые Cyprinidae обитают в озере и образовали там 10 эндемичных видов, представители семейства ограничены ассоциированными водоёмами Лишь Chelaethiops minutus вошёл в состав пелагической ЦС, где он стал единственным потребителем специфического ресурса — насекомых, снесённых в озеро с берега. Интересным представителем зоопланктона пелагиали озера является ручейник Limnoecetis tanganicae. Его крылья редуцированы и он передвигается по поверхностной плёнке, являясь полным аналогом байкальского ручейника Thamastesdipterus. L. tanganicae в массе собирается на свет при ловле пелагической рыбы.
Некоторое развитие получили в озере обитатели пелагиали, живущие только в гиполимнионе (не поднимающиеся в эпилимнион). К таковым можно отнести по крайней мере 10 видов цихлид. Для всех характерно слабо развитие шипов, сильное развитие глаз, мелкие размеры (до 20 см, обычно менее 10 см длиной) и хорошее развитие органов боковой линии. Все они зоопланктофаги.
Как явствует из батиметрической карты, чтобы достичь глубин в 1 км, во многих местах достаточно отойти на лодке от берега на это же расстояние. На столь крутых склонах невозможно отложение осадков, поэтому большая часть дна в пределах аэробной зоны занята выходами скальных пород. С другой стороны, плоский или с малым наклоном шельф также занимает значительную площадь: только на южной оконечности озера более 1000 кв. км. Бенталь озера (если не считать ассоциированных с озером водоёмов) гетерогенна: эпилимнион и гиполимнион населены сильно различающейся фауной бентоса и придонных рыб, различия эти нередко родовые. Имеющиеся данные по составу биоты разных частей мелководий озера не позволяют разделить их на разные биогеографические регионы, поскольку все они могут быть объяснены различием в неравномерном распределении разных биотопов в разных частях озера (Coulter, 1991), в противоположность тому, что утверждал Я.И. Старобогатов (1971: 231).
Итоговая схема биогеографического районирования озера Танганьика следующая: (1) прибрежный район (ассоциированные водоёмы: мелководные заливы, дельты рек и т. п.), (2) пелагиаль открытого озера, (3) мелководный бентический район, (4) глубоководный бентический район. Прибрежный район является частью окружающего озеро биогеографического района, тогда как остальные следует рассматривать как части другого биогеографического региона высокого ранга (вероятно, подобласти), занимающего большую часть озера. При этом все бентические районы следует противопоставить пелагическому, что можно отразить, присвоив бентали и пелагиали ранги провинций.