БИОЛОГИЯ пособие для поступающих в вузы
Том І биология, клетки, генетика и онтогенез, зоология - 2018 год
Пять миллиардов лет назад, до появления жизни на Земле, вся свободная энергия, щедро изливаемая на нашу планету Солнцем, быстро рассеивалась в виде бесполезного тепла и излучалась в космическое пространство. Позднее возникли мельчайшие системы, способные захватывать часть свободной энергии и использовать эту энергию для поддержания своей организации, а также для ее распространения. Таким образом, живая материя не стоит особняком от физического мира, а составляет неотделимую ее часть.
Все живые организмы, обитающие на Земле, представляют собой открытые системы, способные активно организовывать поступление энергии и вещества извне. Энергия необходима для осуществления жизненно важных процессов, но прежде всего — для химического синтеза веществ, используемых для построения и восстановления структур клетки и организма.
Для синтеза компонентов организма необходимо потребление извне химических элементов, используемых в качестве строительных блоков. Главным структурным элементом органических молекул является углерод. В зависимости от источников углерода живые организмы делят на две большие группы: автотрофы, использующие неорганический источник углерода (диоксид углерода), и гетеротрофы, использующие органические источники углерода. Автотрофы самостоятельно синтезируют простые органические соединения, используя для этого неорганические вещества и энергию солнечного излучения (при фотосинтезе) либо энергию окисления неорганических веществ (при хемосинтезе). Гетеротрофы получают простые органические вещества в процессе пищеварения, когда под влиянием пищеварительных ферментов сложные органические компоненты пищи расщепляются на более простые. Однако ни сами органические вещества, ни продукты их расщепления не могут непосредственно служить “топливом” для клеточных процессов. Роль такого универсального топлива играет АТФ (аденозинтрифосфат). Синтез АТФ, этого высококалорийного клеточного топлива, происходит в результате дыхательного обмена — системы множества строго регулируемых химических реакций. Дыхательный обмен поставляет не только энергию, но и строительные материалы. Многие продукты расщепления углеводов, жиров и белков используются для синтеза разнообразных соединений, из которых состоит клетка, в том числе и для синтеза новых углеводов, жиров и белков. Именно это тесное переплетение процессов утилизации веществ для производства энергии и для производства других веществ придает феномену регуляции клеточного метаболизма его уникальный, неповторимый характер. Во всех этих процессах происходит преобразование одного вида энергии в другой.
В общем виде поток энергии можно представить следующим образом.
1. Энергия Солнца —> автотрофы —> органические вещества —> АТФ —> различные формы работы.
2. Энергия Солнца —> автотрофы —> органические веществ —> гетеротрофы —> АТФ —> различные формы работы.
Существуют два типа клеточных механизмов, обеспечивающих поток энергии.
1. Процесс приобретения энергии и вещества живыми организмами называется питанием.
2. Процесс, с помощью которого живые организмы высвобождают энергию из богатых ею веществ, полученных с нищей, называется дыханием.
Автотрофное питание
Автотрофы способны к самостоятельному синтезу необходимых для них органических соединений за счет СО2, воды и энергии света (фотосинтез) или энергии, выделившейся при окислении неорганических соединений (хемосинтез).
Организмы, которые синтезируют все необходимые им органические вещества за счет энергии света, называются фототрофами (это все зеленые растения). Процесс утилизации энергии света называется фотосинтезом. Фотосинтетическими органоидами клеток являются хлоропласты. Мембранная система хлоропластов — это то место, где протекают световые реакции фотосинтеза. В мембранах находится хлорофилл и другие пигменты, ферменты и переносчики электронов.
Структурной и функциональной единицей хлоропластов является тилакоид, имеющий форму плоских мешочков. Тилакоиды местами уложены в стопки — граны. На мембранах тилакоидов имеются частицы двух типов. Такие частицы называются квантосомами. Более мелкие частицы составляют фотосистему 1 (ФС-1), более крупные — фотосистему 2 (ФС-2).
В центре квантосомы находится липопротеидный комплекс, а на внутренней ее стороне — молекулы хлорофилла, каротина и фосфолипида.
Хлорофилл а с максимумом поглощения Р 690 нм и максимумом поглощения Р 700 нм образуют энергетические ловушки и являются основными пигментами, а такие как хл а Р 680 нм, Р 670 нм или хл в, хл с, хл д — вспомогательными. Пигменты улавливают квант света и передают возбужденный электрон (е) к реакционному центру.
Каждая квантосома содержит 200 молекул хлорофилла, но только 10 из них связаны с ферментативным комплексом. В ферментативные комплексы входят цитохромы — это белки, связанные с гемом. Без цитохромов хлорофилл не может передавать возбужденный электрон.
Фотосинтез можно подразделить на две фазы.
1. Световая фаза, которая включает в себя:
а) циклическое фосфорилирование;
б) нециклическое фосфорилирование;
в) фотолиз воды.
2. Темновая фаза.
Световая фаза. При циклическом фосфорилировании квант света ударяет в хлорофилл и выбивает из магния электрон, который тут же захватывается ферментативным комплексом. При этом задействована только фотосистема 1 (ФС-1). Возбужденный электрон проскакивает цитохромы и вновь возвращается на свою орбиту в хлорофилл, но при этом энергия света, которую получил электрон, переводится на цитохромах в энергию химических связей АТФ. Этот механизм, являющийся самым примитивным, имеет место у фотосинтезируемых бактерий:
![]()
АТФ — соединение нестойкое, а в качестве более стойкого акцентора выступает вещество НАДФ. Эта кумуляция (накопление) энергии происходит при нециклическом фосфорилировании. Параллельно с этим процессом происходит и фотолиз воды.
Нециклическое фосфорилирование и фотолиз воды. Нециклическое фосфорилирование наблюдается у растений, имеющих ФС-1 и ФС-2. Квант света выбивает электроны из ФС-1 и ФС-2. Возбужденный электрон из ФС-2 переносится на момекулу акцептора (Q). Молекула хлорофилла ФС-2, потерявшим свои электроны, может заменить их электронами молекуkы воды. Светозависимое окислительное расщепление молекул воды называют фотолизом. Ферменты фотолиза локализованы на внутренней мембране тилакоидов.

В ФС-1 энергия света передает электроны на акцептор P 430, представляющий собой железосерный белок. Далее электроны передаются следующему переносчику — ферредоксину, отдающему свои электроны коферменту НАДФ, который восстанавливается до НАДФ⋅ Н2. Электроны молекулы Р 700 (ФС-1) замещаются электронами ФС-2, поступающими по электронно-транспортной цепи от акцептора Q. При этом происходит фотофосфорилирование с образованием АТФ (рис. 1.17).
Рис. 1.17. Нециклический поток электронов и фотофосфорилирование
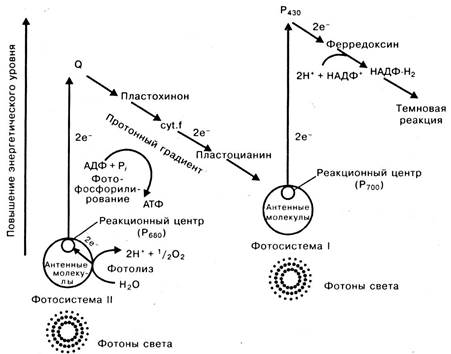
Общее уравнение нециклического фосфорилирования:
Н2O + НАДФ + 2АДФ + 2Ф —> 1/2О2 + НАДФ Н2 + 2АТФ.
В дальнейшем НАДФ⋅ Н2 переходит в строму хлоропласта, где участвует в синтезе углеводов. Для этих реакций свет не нужен, и поэтому они называются темновыми.
Темновая фаза. В темновой фазе происходит перевод активного Н с НАДФ⋅ Н2 на компактный акцептор — углерод, т. е. в темновой фазе синтезируются углеводы. В строму хлоропласта поступает НАДФ⋅ Н2, СО2 и АТФ. Процесс синтеза углеводов происходит циклично и называется циклом Кальвина (рис. 1.18).
Рис. 1.18. Общий итог цикла Кальвина

Исходным (или конечным) соединением цикла Кальвина является сахар — рибулозо-1,5-бифосфат (РБФ). Процесс начинается, когда СО2 входит в цикл и фиксируется на РБФ. Шесть молекул РБФ реагируют с шестью молекулами СО2, образуя 12 молекул 3-фосфоглицерата, которые превращаются в 12 молекул глицероальдегид-3-фосфата. Десять из них перераспределяются с образованием шести пятиуглеродных молекул РБФ. Оставшиеся две молекулы глицероальде- гид-3-фосфата представляют собой “чистый выход” цикла Кальвина. В течение каждого оборота цикла одна молекула СО2 восстанавливается, а молекула РБФ регенерируется. Шесть оборотов цикла с поглощением шести С необходимо совершить, чтобы образовался шестиуглеродный сахар — глюкоза. Суммарное уравнение темновой фазы:
6СО2 + НАДФ⋅ Н 2 + 18АТФ —> С6Н12О6 + 12НАДФ + 18АДФ + 18Ф + 6Н2О.
Помимо углеводов при фотосинтезе образуются промежуточные продукты, например, фосфоглицериновая кислота, которая вступает на путь гликолиза, превращается в ацетильную группу, присоединяется к коферменту А и образует ацетилкофермент А. Из последнего синтезируются жирные кислоты — как в цитоплазме, так и в хлоропластах.
Высшие растения способны синтезировать все необходимые им аминокислоты. Для этого им нужен источник азота или нитратов, а источником углерода служит фосфоглицериновая кислота.
Таким образом, все органические вещества, включающие в себя углерод в качестве акцептора возбужденного электрона, являются источниками энергии.
Кроме фотосинтеза существует еще одна форма автотрофной ассимиляции — хемосинтез, наблюдаемый у некоторых бактерий. Такие бактерии называюется хемосинтетиками.Хемосинтез — это способность хемосинтетиков при окислении неорганических веществ использовать выделившуюся энергию на образование органических веществ, идущих на построение клеток своего тела.
Хемосинтетики в отличие от фототрофов, используют в процессе синтеза не солнечный свет, а энергию экзотермических реакций.
Открытие бактериального хемосинтеза принадлежит С. Н. Виноградскому (1856—1953). Он впервые доказал суще
ствование в природе автотрофных незеленых организмов — серо-, железо- и нитрифицирующих бактерий и описал сущность процесса хемосинтеза.
Нитрифицирующие бактерии, широко распространенные в почве и в различных водоемах, добывают энергию путем окисления аммиака до азотистой кислоты, играя при этом важную роль в круговороте азота в природе. Аммиак, образующийся в почве и водоемах при гниении белков, окисляется нитрифицирующимися бактериями, которым С. Н. Виноградский дал название нитросомонас:
2NН3 + 3O2 ⇔ 2НNO2 + 2Н2O + 158 ккал.
Дальнейшее окисление НNO2 осуществляется другой группой нитрифицирующих микроорганизмов, которые С. Н. Виноградский назвал нитробактером:
2НNO2 + О2⇔ 2HNO3 + 38 ккал.
НNO2 реагирует с минеральными соединениями почвы, превращаясь в соли НNO3.
В водоемах, вода которых содержит Н2S, живут бесцветные серобактерии. Энергию, необходимую для синтеза органических соединений, из СО2 они получают, окисляя Н2S:
2Н2S + O2 = 2Н2O + 2S.
При недостатке Н2S серобактерии производят дальнейшее окисление:
2S + 3O2 + 2Н2O = 2Н2SO4 + 115 ккал.
Колоссальное количество серобактерий имеется в Черном море.
Железобактерии окисляют закисные соединения железа в окисные, участвуя в круговороте железа в природе:
4FеСO3 + O2 + 6Н2O = 4Fе(ОН)3 + 4СO2 + 81 ккал.
Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Известны также бактерии-хемосинтетики, способные окислять метан, окись углерода, водород.
Таким образом, хемосинтетики являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения (углеводы, липиды, белки и др.), а не берут их в готовом виде, как гетеротрофы.