БИОЛОГИЯ пособие для поступающих в вузы
Том ІІ ботаника, анатомия и физиология, эволюция и экология - 2018 год
Эволюция как реальность (признаки и доказательства). Понятия вида и популяции - ЭВОЛЮЦИЯ ОРГАНИЗМОВ - ЭВОЛЮЦИЯ И ЭКОЛОГИЯ
Часто интуитивно мы считаем, что вид — это группа очень сходных между собой организмов, например, людей, лошадей или морковок. Научное определение вида неоднократно изменялось, и в настоящее время наиболее широко распространенным является следующее: вид — это группа морфологически сходных организмов, имеющих общее происхождение и потенциально способных к скрещиванию между собой в естественных условиях. Особи, из которых слагается вид, не всегда живут в непосредственном соседстве друг с другом. Обычно они образуют более или менее изолированные группы, называемые популяциями. Изучая процесс эволюции, мы обычно рассматриваем составляющие вид популяции. Популяция — это сообщество скрещивающихся между собой организмов. Особи скрещиваются друг с другом в пределах данной популяции либо могут покидать свою популяцию и скрещиваться с особями, принадлежащими другой популяции.
Критерии вида.
1. Морфологический критерий свидетельствует о морфологической изоляции вида. Он является одним из самых ранних видовых критериев. Им пользовались систематики всех поколений, начиная с Линнея. Он наиболее прост и удобен в практической работе. На его основе составлены определительные таблицы как в ботанике, так и в зоологии.
Ограниченность морфологического критерия выступает особенно отчетливо на примере так называемых видов-двойников, или биологических видов. Виды- двойники можно определить, как морфологически сходные или идентичные, репродуктивно изолированные природные популяции.
2. Физиолого-биохимический критерий. В основе этого критерия заложено следующее понятие: виды различны, потому что различны их химические структуры. Особенности вида передаются от поколения к поколению, так как в гаметах закодированы процессы синтеза определенных, специфических белков. Отсюда следует, что для биохимической оценки видовой специфики особый интерес представляет изучение особенностей ДНК вида. Изучение видовой специфики ДНК естественно подводит нас к следующему видовому критерию.
3. Генетический критерий. В основе его лежат два положения. Во-первых, виды различаются кариотипами, т. е. совокупностью признаков хромосомного комплекса. Во-вторых, особи разных видов в естественных условиях, как правило, не скрещиваются. Препятствием для скрещивания являются различные изолирующие механизмы (физиологические, морфологические, экологические), вырабатывающиеся в результате эволюционного процесса. Однако для большинства животных изолирующие механизмы, предотвращающие скрещивание близкородственных видов, до конца не изучены. Определение вида, которое возникает из такой теоретической концепции, гласит: виды — это группы скрещивающихся естественных популяций, репродуктивно изолированные от других таких групп.
Однако существуют виды, которые не изолированы друг от друга барьером стерильности. Например, кряква и шилохвость, два разных вида уток, в неволе способны скрещиваться друг с другом и давать многочисленное потомство. Очевидно, разобщенность этих двух видов поддерживается какими-то другими факторами.
4. Эколого-географический критерий. Используется с целью установления закономерностей географического распространения биологических форм и особенностей их взаимоотношения с факторами внешней среды. Каждый вид занимает определенную область распространения, т. е. ареал. Ареалы видов одного рода могут быть полностью изолированными либо совпадать частично или полностью. Виды, имеющие изолированные ареалы, называются аллопатрическими. Виды, у которых ареалы перекрываются или совпадают, именуются симпатрическими. Видовая специфика каждого из них вскрывается в результате их экологических особенностей. Например, отсутствие гибридов между органическими формами в местах перекрывания ареала.
Иерархическая классификация организмов (таксономия)
Нетрудно видеть, что все рассмотренные нами критерии дополняют друг друга, помогая вскрывать специфику вида. Поэтому в практике описания видов животных и растений они используются все вместе, в зависимости от того, насколько изучены те или иные стороны морфологии, физиологии, генетики и экологии изучаемой формы.
Виды, совпадающие по многим существенным признакам, но все же репродуктивно изолированные друг от друга, объединяются в роды. По мере ступенчатого расширения критериев сходства близкие между собой роды соединяются в семейства, семейства — в отряды, отряды — в классы, классы — в типы (рис. 6.32). Иерархическое соподчинение этих таксонов должно указывать на степень филогенетического родства. В построенной таким образом естественной системе объединение видов в таксон высшего ранга говорит о наличии у них общей исходной формы в определенное геологическое время, причем чем дальше от современности момент рождения родственных форм, тем выше должен быть ранг охватывающей виды систематической категории.
Рис. 6.32. Диаграмма взаимоотношений между систематическими категориями
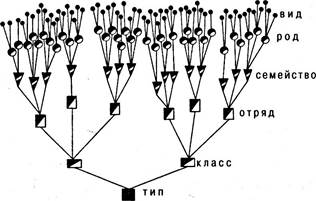
Такие категории, как особи, скрещивающиеся популяции, виды, — это естественные, реально существующие объекты, тогда как таксоны более высокого ранга представляют собой абстрактные логические категории. Каждая из них — это класс объектов, в который входят все категории более низкого ранга, имеющие определенные общие признаки. Добавляя к вышеназванным категориям префиксы под- и над- (в систематике животных), можно создавать промежуточные категории, например, подвиды, надсемейства и т. п.
Сама возможность построения иерархической классификации организмов (таксономия) по различным уровням сходства объяснима только тем, что категории этой системы отражают в той или иной степени близкое или отдаленное филогенетическое “кровное родство”. Именно в их выявлении состоит смысл таксономических исследований.
Итак, таксономия, или систематика, дает весьма ценные доказательства в пользу эволюционного учения.
1. Независимо от эволюционного учения систематика обнаруживает родственные связи между организмами.
2. Независимо от эволюционного учения систематика приходит к выводу о необходимости изображать отношения между естественными группами организмов в виде генеалогического древа.
3. Систематика накапливает ценный материал по вопросам изменчивости и изменяемости видов и дает основания считать справедливым взгляд, согласно которому виды эволюционируют.
Доказательства эволюции из области сравнительной анатомии
Сравнительная анатомия изучает строение животных (или растений), в результате чего удается установить черты сходства и различия и определить степень родства между ныне живущими организмами.
В общей форме естественное сходство выражается в том, что у сравниваемых форм наблюдается сходный общий план организации, так называемое единство плана строения. В узком смысле закон единства плана строения применим к отношениям, существующим в пределах каждого отдельного типа животного царства. Например, сравнивая скелеты позвоночных животных, легко установить в общей организации, расположении, числе и форме частей этих скелетов взаимное сходство (рис. 6.33). Факт замечательного общего сходства в строении позвоночных естественнее всего объясняется предположением, что все эти формы связаны узами родства.
Рис. 6.33. Единство плана строения: 1 — скелет человека; 2 — скелет шимпанзе; 3 — скелет лягушки
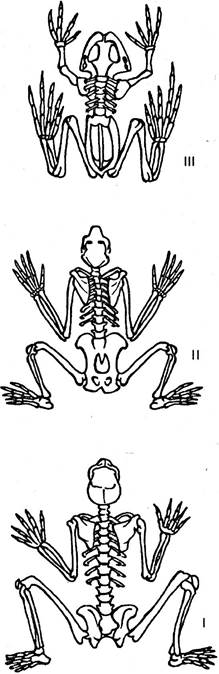
Возникает, однако, вопрос: возможно ли более широкое толкование закона единства плана строения? Можно ли, например, применить это понятие не только к отношениям, существующим в пределах одного класса, но и к отношениям, существующим между типами? На этот вопрос в ряде случаев можно дать положительный ответ, в особенности если мы будем искать единство плана строения не только среди взрослых, но и личиночных форм. При такой оговорке можно легко показать замечательное сходство, например, между личиночными формами кольчатых червей и некоторых моллюсков, что позволяет ставить вопрос о родстве между этими двумя типами.
В ряде случаев сходен не только общий план строения сравниваемых форм, но и детали их организации. Это выражается, в частности, в установлении гомологии органов, имеющей первостепенное значение для выяснения филогенетического родства между организациями.
Гомологичные органы — это такие органы разных животных (растений), которые имеют сходное строение, одинаковое положение в теле и развиваются из тех же самых эмбриональных закладок. Гомологичные органы могут выполнять у разных животных одинаковые или разные функции. Например, иглы ежа гомологичны волосам других млекопитающих. Усики гороха, служащие для прикрепления его стебля к другим растениям, являются гомологами листа.
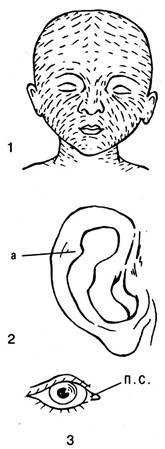
Рудиментарные органы. Особым видом гомологичных органов являются рудименты (зачаток). Это органы, утерявшие в филогенезе свое значение и функцию и оставшиеся в виде недоразвитых образований. Поэтому точнее было бы назвать их не зачаточными, а остаточными. В теле каждого животного можно встретить много рудиментов. Немало рудиментов имеется и в теле человека (рис. 6.34).
Рис. 6.34. Рудиментарные органы человека: 1 — волосяной покров на голове пятимесячного эмбриона; 2 — ушная раковина с Дарвиновым бугорком (а); 3 — полулунная складка (п. с.)
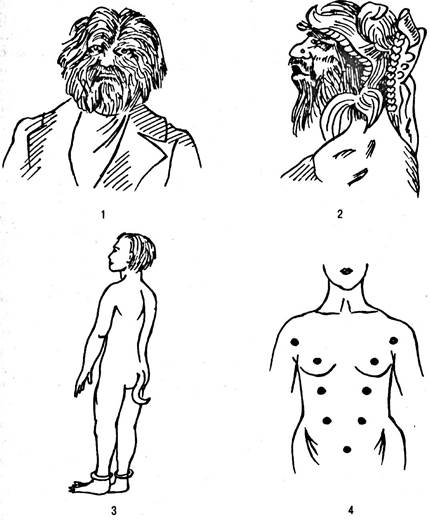
Атавизм — это появление признаков, присущих далеким предкам и в норме не встречающихся у современных форм. Естественно, что такие признаки говорят о филогенетическом родстве между различными организмами: теми, у кого такие признаки отсутствуют как норма, и теми, у кого они проявляются лишь как атавизм. Примеры атавизма у человека — шейная фистула (образование, гомологичное жаберной щели), хвостовой придаток, обильная волосатость всего тела, наличие более двух молочных желез и т. д. (рис. 6.35).
Рис. 6.35. Развитие атавистических признаков у людей: 1—2 — волосатые люди: Адриан Евтихеев (1) и мексиканская танцовщица Юлия Пастрана (2); 3 — десятилетний мальчик из Индии с мягким хвостом; 4 — случай многососковости
Аналогичные органы — это органы, выполняющие одинаковую функцию, но не имеющие единства строения и происхождения. Например, у рыб органы водного дыхания — жабры. У ракообразных, органами дыхании тоже служат жабры, но они находятся в конечностях. Тот факт, что эти органы развиваются различным путем, указывает, что морфологическое сходство между ними чисто внешнее, не отражающее родства между организмами.
Конвергенция. Во многих случаях, когда неродственные организмы живут в одинаковых условиях внешней среды и ведут сходный образ жизни, у них под влиянием естественного отбора возникают аналогичные органы и сходные формы тела. Такое сходство можно видеть, сравнивая быстро плавающих морских хищников — акулу (класс рыб), ихтиозавра (класс пресмыкающихся) и дельфина (класс млекопитающих).
Учение о гомологии и аналогии органов уточнило методы систематики (таксономии), позволило избегать ошибочного объединения животных в систематические группы на основе лишь внешнего сходства. Классификация, построенная на эволюционном принципе, доказывает, что все систематические группы — типы, классы, отряды — связаны друг с другом общностью происхождения. Свидетельством этому служат так называемые сборные формы, т. е. животные, сочетающие признаки разных систематических групп. Так, ланцетник связывает позвоночных с беспозвоночными. Кистеперые рыбы обладают и жабрами и легкими, связывая тем самым рыб с амфибиями.
Доказательства эволюции на базе эмбриологии
Сравнительный метод исследования фактов эмбрионального развития дает ценные свидетельства в пользу эволюционного учения. Между зародышами животных наблюдается необычайное сходство, с предельной ясностью указывающее на “кровное родство”.
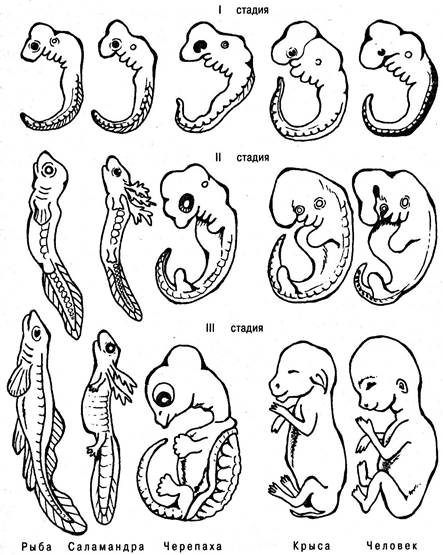
На основании сравнительно-эмбриологических исследований К. Бэр сформулировал закон зародышевого сходства, согласно которому черты, общие для большой группы родственных животных, проявляются раньше частных признаков. По Бэру, в зародышевом развитии животных наиболее рано формируются признаки, отражающие принадлежность к определенному классу, затем отряду, роду и, наконец, виду. Закладка органов, их положение, форма, способы развития, отношение друг к другу необычайно сходны, напри мер, у зародыша рыбы, саламандры и черепахи, у эмбриона крысы и человека (рис. 6.36). В дальнейшем развитии зародышей позвоночных отмеченное сходство ослабевает. По степенно зародыши как бы “расходятся” и явственно приобретают черты, свойственные уже не только позвоночным вообще, но и определенным классам, к которым они принадлежат.
Рис. 6.36. Эмбрионы позвоночных животных на ранних стадиях развития (I стадия) сходны друг с другом, на более поздних (II и III стадии) похожи на филогенетически родственные формы
Сопоставление организации низших и высших животных с их зародышевыми стадиями показывает, что каждое высшее животное проходит в своем индивидуальном развитии этапы, соответствующие более низкоорганизованным животным. Это представление позволило Э. Геккелю в 1866 г. сформулировать основной биогенетический закон, в соответствии с которым онтогенез представляет собой краткое и быстрое повторение филогенеза. Согласно Геккелю, новые признаки, имеющие эволюционное значение, возникают во взрослом состоянии. По мере усложнения организации взрослых форм зародышевое развитие удлиняется за счет включения дополнительных стадий. Этим законом объясняется далеко идущее сходство между эмбриональными стадиями животных. Например, тот факт, что зародыши всех высших животных имеют жаберные карманы в глотке, находит свое наиболее естественное объяснение в утверждении, что все они генетически родственны и произошли в конечном счете от предков, живших в воде.
Тот факт, что все многоклеточные организмы начинают свое индивидуальное развитие с яйца (с одноклеточного состояния) и проходят сходные стадии эмбрионального развития, дает право предлагать и единственно естественное объяснение этого факта, а именно, что многоклеточные организмы произошли от одноклеточных предков, т. е. от одного общего корня.
Начиная с Геккеля, биологи широко использовали биогенетический закон в целях выявления филогении определенных систематических групп. Однако результаты применения этого закона к выявлению закономерностей эволюции не всегда оказывались удовлетворительными. Сам Геккель был принужден констатировать, что в ряде случаев в онтогенезе не происходит рекапитуляции, т. е. повторения пройденного предками пути исторического развития. В этой связи он выделил палингенезы и ценогенезы.
Под палингенезами он понимал повторение филогенетических изменений в индивидуальном развитии.
Ценогенезы, по Геккелю, представляют собой эмбриональные нарушения, происходящие в результате приспособления зародышей или молодых особей к определенным условиям индивидуального развития. Геккель различал два типа ценогенезов — гетеротопии и гетерохронии. При гетеротопии происходит сдвигание местоположения органов; гетерохронии представляют собой нарушения во время закладки органов. Примером гетерохронии служит более ранняя закладка нервной системы и запаздывание в формировании половой системы у высших позвоночных и человека по сравнению с низшими. Примером гетеротопии является закладка легких, представляющих собой видоизменение задней пары жаберных мешков, расположенных по бокам кишечника, на его брюшной стороне. Примером ценогенеза можно считать появление провизорных органов у зародышей наземных позвоночных (амниона, хориона, аллантоиса).
Крупным шагом в развитии эволюционной эмбриологии явилось учение о филэмбриогенезах, созданное А. Н. Северцовым.
Филэмбриогенезами называют новые наследственные свойства, которые возникают в зародышевой жизни животных, сохраняются у взрослых форм и передаются последующим поколениям, включаясь, таким образом, в филогенез. Филэмбриогенезы от ценогенезов отличаются тем, что являются не приспособлениями зародыша, а новыми свойствами, остающимися у взрослых животных.
Согласно Северцову, филэмбриогенезы могут возникнуть на любой стадии эмбрионального развития. Основное положение теории заключается в том, что эволюционные изменения состоят в изменении хода онтогенетического развития. Различают три основных способа филэмбриогенезов: анаболия, девиация, архаллаксис.
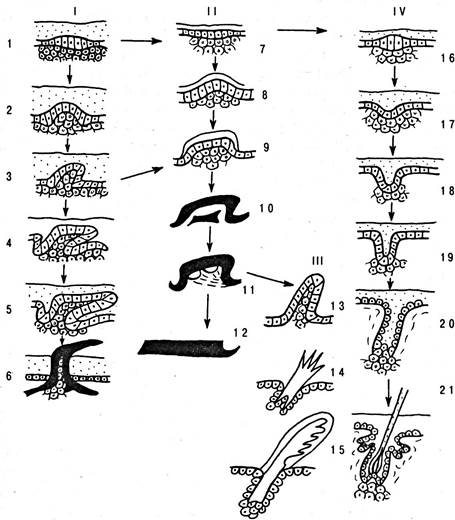
Под анаболией понимают такой способ изменений, который возникает на последних фазах развития. Девиациями хода онтогенеза называют изменения, происходящие на средних фазах морфогенеза. Наконец, к архаллаксисам относят такие изменения, которые происходят в самом начале эмбрионального развития органа и поэтому резко изменяют весь ход морфогенеза. Примером всех трех видов филэмбриогенезов могут служить изменения в развитии придатков кожи пресмыкающихся, птиц и млекопитающих по сравнению с таковыми у хрящевых рыб, древние виды которых были предками наземных позвоночных (рис. 6.37).
Рис. 6.37. Схема филогенетического развития костных (I) и роговых (II) чешуй, перьев (III) и волос (IV) позвоночных: 1 — 6 — развитие плакоидной чешуи акулы; 7—12 — развитие чешуй ящерицы. Ход их развития изменен на средних стадиях (девиация); 13 — 15 — развитие пера. Сперва развитие пера повторяет пути развития чешуи пресмыкающихся (7 — 11), но на последних стадиях (11) происходит изменение (анаболия) и развитие пера идет по новому пути (13 — 15); 16 — 21 — развитие волоса. С наиболее ранних стадий направление развития волоса уклоняется от направлений развития прочих кожных образований; повторения нет, наблюдается архаллаксис
Доказательства эволюции из области палеонтологии
Изучение процесса эволюции на основании палеонтологических материалов привело к формулировке ряда законов, часть которых в значительной мере правильно обобщает наблюдения, другие же требуют некоторых поправок. Приведем наиболее известные законы.
1. Закон неспециализированности. Согласно этому закону, к эволюционному развитию способны малоспециализированные формы; прогрессирующая специализация отдельных ветвей ведет к вымиранию соответствующей группы организмов. Причину этого вымирания можно объяснить прогрессивно уменьшающейся изменчивостью специализированных форм. Вымирание специализированных ветвей в связи с пониженной изменчивостью не может, однако, считаться универсальной закономерностью, так как даже среди очень крупных специализированных форм, например, бабочек, являющихся конечной филогенетической ветвью насекомых, изменчивость чрезвычайно велика.
2. Закон необратимости эволюции. Согласно этому закону, эволюция организмов, изменившихся в каком-нибудь направлении, никогда не приводит к появлению признаков, характерных для его предков. То есть эволюционный процесс не воз вращается вспять по старому пути. Так, лошадь, утратившая все пальцы, кроме одного, ни при каких обстоятельствах не сможет развить свои рудиментарные пальцы.
На основании остатков организмов, находимых в разных слоях почвы, можно судить об их строении и отчасти об их жизнедеятельности, о периоде жизни Земли, когда эти организмы ее населяли, об их распространении и, что самое глав ное, об их генетической связи друг с другом и с современными живыми существами.
Чтобы убедиться в этом, попытаемся рассмотреть общие направления эволюционного процесса и смену форм во времени (табл. 6.1).
Таблица 6.1
|
Эра |
Периоды |
Характерные группы организмов |
|
Докембрийская |
Плохая сохранность палеонтологических документов. Случайные находки. Кольчатые черви (аннелиды), ракообразные (трилобиты). |
|
|
Палеозойская |
Кембрийский |
Среди простейших уже имеются и современные роды. Кишечнополостные: еще нет настоящих кораллов. Черви', высокодифференцированные аннелиды. Членистоногие'. низшие ракообразные. Иглокожие: древние формы. |
|
Силурийский |
Кишечнополостные: появились настоящие кораллы. Иглокожие: древние группы в расцвете. Еще мало морских звезд и ежей. Моллюски:бросается в глаза множество примитивных головоногих. Членистоногие: трилобиты и ракоскорпионы. Первые членистоногие, дышащие воздухом, в том числе древние скорпионы. Позвоночные: низшие формы, так называемые бесчелюстные, в том числе так называемые панцирные рыбы. Настоящие рыбы: древние акуловые, двоякодышащие, кистеперые. Растения: водоросли, некоторые папоротники. |
|
|
Девонский |
Кишечнополостные: многие древние формы в упадке. Кораллы многочисленны. Иглокожие: древние (силурийские) морские лилии. Морских ежей еще мало. Моллюски: среди головоногих все еще господствуют примитивные наутилиды. Позвоночные: древние рыбообразные панцирные формы достигают расцвета. Рыбы: акуловые, химеры, древние кистеперые, примитивные осетровые, двоякодышащие. Земноводные:появились стегоцефалы. Растения: первые многочисленные наземные формы, хвощи, папоротники и др. |
|
|
Каменноугольный |
Кишечнополостные: много древних кораллов, ныне не существующих. Иглокожие: многочисленные морские лилии, древние ежи, ныне не существующие. Моллюски: среди брюхоногих моллюсков много легочных (наземных) форм. Членистоногие: многоножки, пауки с членистым брюшком, много насекомых, в том числе представители отрядов: бескрылых, прямокрылых, сетчатокрылых, полужесткокрылых (клопы). Ракоскорпионы в упадке. Трилобиты в упадке. Позвоночные: двоякодышащие и |
|
|
Палеозойская |
кистеперые рыбы сокращаются в числе. Большим числом форм представлены примитивные земноводные — стегоцефалы. И конце периода появляются первые пресмыкающиеся. Растения: папоротники, хвощи, некоторые голосеменные. |
|
|
Пермский |
Моллюски: много аммонитов и наутилид. Членистоногие: трилобиты вымирают. Позвоноч ные: прежние группы рыб, много настоящих акул. Земноводные: многочисленные и разнообразные стегоцефалы. Пресмыкающиеся. примитивные группы. Растения: развитие хвойных. |
|
|
Мезозойская |
Триасовый |
Простейшие: много современных родов. Кишечнополостные: среди кораллов смена дрен них новыми, шестилучевыми. Иглокожие: среди морских лилий многие вымерли, но все же эта группа господствует. Морские ежи многочисленнее, древние вымерли, появились новые формы. Моллюски: в расцвете, много новых групп. Мезозойские брюхоногие моллюски вымирают, наутилиды сокращаются в числе. Аммонитов много, н том числе новые формы. Белемниты еще редки. Членистоногие: трилобиты исчезли, появляются десятиногие раки. Позвоночные: появляются костистые рыбы. Земноводные: последние стегоцефалы. Пресмыкающиеся: динозавры, крокодилы, черепахи, плезиозавры, ихтиозавры и др. Появляются млекопитающие: примитивные группы сборного и переходного типов. Растения: среди хвойных — араукария и сосна. |
|
Юрский |
Кишечнополостные: пышный расцвет рифообразующих кораллов. Иглокожие: все еще много морских лилий. Морские ежи еще многочисленнее, чем в триасе. Моллюски: множество новых форм. Кораблики в упадке и представлены одним родом — Наутилус (кораблик). Белемниты в расцвете, новые аммониты. Членистоногие: среди насекомых первые представители двукрылых и перепончатокрылых. Позвоночные. Рыбы: характерны примитивные костистые. Пресмыкающиеся: в расцвете диплодок, стегозавр, игуанодон, черепахи, крокодилы, ихтиозавры, плезиозавры и др. Появляются птицы: археоптерикс, археоникс. Млекопитающие: еще далеко до расцвета, мелкие примитивные формы. Растения: хвощи, папоротники, хвойные. |
|
|
Мезозойская |
Меловой |
Иглокожие: морские лилии в упадке. Первое место переходит к морским ежам. Моллюски: множество аммонитов и белемнитов. Наутилус (кораблик). Членистоногие: много десятиногих раков. Рыбы: массовое развитие костистых рыб (расцвет). Земноводные: появляются формы, близкие к современным. Пресмыкающиеся в расцвете. Появляются настоящие ящерицы и крокодилы. Ихтиозавры вымирают. Зубастые птицы. Млекопитающие: те же древние мелкие формы, но появляются сумчатые и неполнозубые. Растения: еще в нижнем мелу появляются однодольные и двудольные, в том числе ива, тополь, дуб, береза, эвкалипт и др. |
|
Кайнозойская |
Третичный |
Вымирают новые формы мелового периода. Иглокожие: много современных родов. Моллюски: белемниты и аммониты приходят в упадок и вымирают. Позвоночные: высшие костистые рыбы. Земноводные: близки современным. Вымирают все древние пресмыкающиеся, и остаются гаттерии, ящерицы, крокодилы и черепахи. Впервые появляются змеи. Среди птиц много близких современным. Происходит закладка современных отрядов млекопитающих. |
|
Четвертичный |
Современные формы животного и растительного мира. Человек. |