Биология в экзаменационных вопросах и ответах для абитуриентов, репетиторов, учителей
Каковы особенности строения, размножения и жизнедеятельности растений в связи с выходом на сушу - Растения - МНОГООБРАЗИЕ ОРГАНИЧЕСКОГО МИРА
Существует предположение, что растения произошли от древних зеленых водорослей. Для такого предположения имеются веские основания: 1) у водорослей и растений главный фотосинтезирующий пигмент — хлорофилл а и каротиноиды; 2) основной запасной углевод — крахмал, который откладывается в хлоропластах, а не в цитоплазме, как у других фотосинтезирующих эукариот; 3) целлюлоза является важнейшим компонентом их клеточной стенки; 4) у водорослей и некоторых растений (мхи) в матриксе хлоропласта имеются особые включения — пиреноиды; 5) есть сходство в клеточном делении растений и некоторых водорослей.
Растения появились на суше примерно 430 млн лет назад в виде небольших по размеру и примитивных по строению риниофитов и псилофитов. В дальнейшем их эволюция неразрывно связана с постепенным завоеванием суши. Оказавшись в совершенно новой воздушной среде, они постепенно адаптировались к необычной обстановке и на протяжении многих миллионов лет дали большое разнообразие наземных растений.
Самой большой трудностью, которую надо было преодолеть первым наземным растениям, была проблема обезвоживания. Как известно, у водорослей мужские гаметы, участвующие в половом размножении, имеют возможность приблизиться к яйцеклеткам только в водной среде. На суше такая возможность крайне ограничена. Одним из ключевых событий раннего этапа выхода растений на сушу явилось появление спор с прочными оболочками, позволяющими переносить засушливые условия. В процессе дальнейшего приспособления к наземным условиям существования у растений сформировались вегетативные органы — корень, стебель и лист, как результат разделения функций между различными участками тела. Корни обеспечивают закрепление растений в субстрате и водно-минеральное питание, листья — фотосинтез, стебли — транспорт веществ (восходящий и нисходящий токи). Расчленение тела растений на органы явилось следствием развития у них разнообразных тканей — покровной, проводящей, механической и др.
Развитие у сосудистых растений эффективной проводящей системы, состоящей из ксилемы и флоэмы, решило проблему передвижения воды, минеральных и органических веществ в условиях суши. От высыхания растения защищены покровной тканью — эпидермисом, перидермой или коркой. В свою очередь развитие эпидермиса привело к формированию устьиц — высокоспециализированных образований, регулирующих газообмен и испарение воды растением.
У растений наблюдается усиление механической устойчивости стебля вследствие утолщения клеточной стенки и пропитывания ее лигнином, придающим жесткость целлюлозному остову клеточной оболочки. Многие ученые полагают, что появление способности к синтезу лигнина оказалось поворотным моментом в эволюции наземных растений.
Параллельно шла эволюция органов полового и бесполого размножения. Из одноклеточных, характерных для подавляющего большинства водорослей, антеридии и архегонии становятся многоклеточными, их стенки надежнее защищают гаметы и споры от высыхания.
В жизненном цикле растений наблюдается закономерное чередование полового и бесполого поколений. Это значит, что в ходе жизненного цикла (т.е. цикла от зиготы одного поколения до зиготы следующего поколения) один тип организмов сменяется другим. Схема такого цикла изображена на рис. 44.
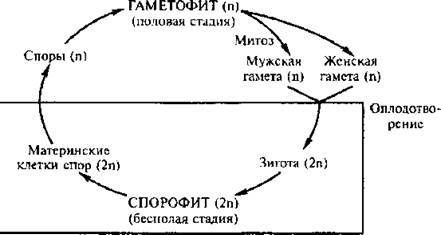
Рисунок 44. Обобщенная схема жизненного цикла растения, у которого наблюдается чередование поколений
Гаплоидное поколение называется гаметофитом, оно способно к половому размножению и образует гаметы. Гаметы формируются в многоклеточных органах полового размножения — антеридиях и архегониях.
Антеридии — небольшие овальные тельца, внешняя стенка которых образована одним, реже — несколькими слоями стерильных клеток. В антеридиях формируются подвижные мужские гаметы — сперматозоиды, которые выходят из созревшего антеридия наружу и активно передвигаются только в воде.
Архегонии — небольшие колбообразные тельца, состоящие из нижней расширенной части, называемой брюшком, и верхней удлиненной — шейки. Снаружи архегонии окружают бесплодные клетки, защищающие их от высыхания. В брюшке находится неподвижная женская гамета — яйцеклетка. Над яйцеклеткой располагается брюшная канальцевая клетка, а внутри шейки — ряд шейковых канальцевых клеток.
При созревании яйцеклетки брюшная и шейковые канальцевые клетки расплываются в слизь, и архегоний вскрывается на верхушке. По каналу шейки, заполненному слизью, сперматозоиды проникают в брюшко, и один из них сливается с яйцеклеткой, т.е. происходит оплодотворение. В результате образуется диплоидная зигота, из которой вырастает диплоидный спорофит. Он называется спорофитом, так как способен к бесполому размножению с образованием гаплоидных спор. Последние дают начало гаметофитному поколению.
Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла.
В отличие от других растений в жизненном цикле моховидных преобладает гаметофит — небольшое, преимущественно листостебельное растение, которое осуществляет функции фотосинтеза, водоснабжения и минерального питания. Спорофит у них развивается из оплодотворенной яйцеклетки внутри архегония и постоянно связан с гаметофитом не только морфологически, но и физиологически, являясь его придатком. Для размножения мхов необходима вода, иначе сперматозоиды не смогут подплыть к архегониям. Кроме того, только в достаточно влажной среде у них лопаются антеридии и высвобождаются сперматозоиды. Поэтому не случайно большинство моховидных приурочены к сырым и затененным местам.
Развитие моховидных по пути возрастания самостоятельности гаметофита и упрощения спорофита привело к эволюционному тупику.
В эволюции сосудистых растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита, который не нуждается для образования и распространения спор в капельножидкой среде. Так, у плауновидных, хвощевидных и папоротниковидных гаметофит представлен маленьким (от нескольких миллиметров до 3 см) заростком, не расчлененным на органы, живущим несколько недель (у плаунов — несколько лет) независимо от спорофита. На заростках в антеридиях развиваются сперматозоиды, которые, плавая в каплях воды, достигают архегония и сливаются с яйцеклеткой. Благодаря крошечным размерам гаметофитов оплодотворение у хвощей, плавунов и папоротников может происходить даже; при ничтожно малых количествах воды в виде капелек росы, тумана и др.
У голо- и покрытосеменных растений гаметофит полностью утратил способность к самостоятельному образу жизни. Все его развитие протекает на спорофите внутри макроспорангия (или семязачатка).
У голосеменных женский гаметофит — многоклеточный гаплоидный эндосперм с двумя (у сосны) или несколькими (у других голосеменных) архегониями; у покрытосеменных он редуцирован обычно до семи клеток, архегониев не имеет и называется зародышевым мешком. В зародышевом мешке образуются яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид, вторичное (диплоидное) ядро и клетки-антиподы.
Мужской гаметофит семенных растений развивается из микроспоры и представляет собой пыльцевое зерно (пыльцу). Оно содержит клетку, прорастающую в пыльцевую трубку, и еще генеративную, образующую два спермия. При этом впервые в эволюции растений процесс оплодотворения становится независимым от наличия капельножидкой среды; спермин доставляются к яйцеклеткам пыльцевой трубкой, что является важнейшим приспособлением к наземному образу жизни.