Биология - Универсальный справочник школьника
ГЕНЕТИКА
Генетика как наука
Генетика — это наука, изучающая закономерности наследственности и изменчивости и методы управления ими.
Задачами генетики на современном этапе являются исследование качественных и количественных характеристик наследственного материала, анализ структуры и функционирования генотипа, расшифровка тонкой структуры гена и методов регуляции генной активности, поиск генов, вызывающих развитие наследственных болезней человека и методов их «исправления», создание нового поколения лекарственных препаратов по типу ДНК-вакцин, конструирование с помощью средств генной и клеточной инженерии организмов с новыми свойствами, которые могли бы производить необходимые человеку лекарственные препараты и продукты питания, а также полная расшифровка генома человека.
Наследственность — это способность организмов передавать свои признаки и свойства в ряду поколений.
Изменчивость — свойство организмов приобретать новые признаки в течение жизни.
Признаки организмов можно разделить на качественные и количественные. Качественные признаки имеют два-три контрастных проявления, которые называют альтернативными признаками, например, голубой и карий цвет глаз, тогда как количественные (удойность коров, урожайность пшеницы) не имеют четко выраженных различий.
Материальным носителем наследственности является ДНК. У эукариот различают два типа наследственности: генотипическую и цитоплазматическую. Носители генотипической наследственности локализованы в ядре, а носителями цитоплазматической наследственности являются находящиеся в митохондриях и пластидах кольцевые молекулы ДНК. Цитоплазматическая наследственность передается в основном с яйцеклеткой, поэтому называется также материнской.
В митохондриях клеток человека локализовано небольшое количество генов, однако их изменение может оказывать существенное влияние на развитие организма, например, приводить к развитию слепоты или постепенному снижению подвижности. Пластиды играют не менее важную роль в жизни растений. Так, в некоторых участках листа могут присутствовать бесхлорофильные клетки, что приводит, с одной стороны, к снижению продуктивности растения, а с другой — такие пестролистные организмы ценятся в декоративном озеленении. Воспроизводятся такие экземпляры в основном бесполым способом, так как при половом размножении чаще получаются обычные зеленые растения.
Методы генетики:
1. Гибридологический, или метод скрещиваний, заключается в подборе родительских особей и анализе потомства. При этом о генотипе организма судят по фенотипическим проявлениям генов у потомков, полученных при определенной схеме скрещивания.
2. Цитогенетический основан на исследовании кариотипа: числа, формы и величины хромосом организма. Изучение этих особенностей позволяет выявить различные патологии развития.
3. Биохимический позволяет определять содержание различных веществ в организме, в особенности их избыток или недостаток, а также активность целого ряда ферментов.
4. Молекулярно-генетические направлены на выявление вариаций в структуре и расшифровку первичной последовательности нуклеотидов исследуемых участков ДНК. Они позволяют выявить гены наследственных болезней даже у эмбрионов, установить отцовство и т. д.
5. Популяционно-статистический позволяет определить генетический состав популяции, частоту определенных генов и генотипов, генетический груз, а также наметить перспективы развития популяции.
6. Гибридизации соматических клеток в культуре, который позволяет определить локализацию определенных генов в хромосомах при слиянии клеток различных организмов, например, мыши и хомяка, мыши и человека и т. д.
Основные генетические понятия и символика
Ген — это участок молекулы ДНК, или хромосомы, несущий информацию об определенном признаке или свойстве организма. Некоторые гены могут оказывать влияние на проявление сразу нескольких признаков. Такое явление называется плейотропией.
Каждый ген занимает в хромосоме строго определенное место — локус. Так как в соматических клетках большинства эукариотических организмов хромосомы парные (гомологичные), то в каждой из парных хромосом находится по одной копии гена, отвечающего за определенный признак. Такие гены называются аллельными.
Аллельные гены чаще всего существуют в двух вариантах — доминантном и рецессивном. Доминантной называют аллель, которая проявляется вне зависимости от того, какой ген находится в другой хромосоме и подавляет развитие признака, кодируемого рецессивным геном. Доминантные аллели обозначаются обычно прописными буквами латинского алфавита (A, B, Cи др.), а рецессивные — строчными (a, b, c и др.). Рецессивные аллели могут проявляться только в том случае, если они занимают локусы в обеих парных хромосомах.
Организм, у которого в обеих гомологичных хромосомах находятся одинаковые гены, называется гомозиготным по данному гену, или гомозиготой (AA, aa, AABB, aabb и т. д.), а организм, у которого в обеих гомологичных хромосомах находятся разные варианты гена — доминантный и рецессивный — называется гетерозиготным по данному гену, или гетерозиготой (Aa, AaBb и т. д.).
Ряд генов может иметь три и более структурных вариантов, например, группы крови по системе АВО кодируются тремя аллелями — IA, IB, i. Такое явление называется множественным аллелизмом. Однако даже в этом случае каждая хромосома из пары несет только один аллель, то есть все три варианта гена у одного организма представлены быть не могут.
Совокупность генов, характерную для гаплоидного набора хромосом называют геномом, тогда как генотипом — совокупность генов, характерную для диплоидного набора хромосом. Совокупность признаков и свойств организма, которая является результатом взаимодействия генотипа и окружающей среды, называется фенотипом.
Для записи схем скрещиваний чаще всего применяются следующие условные обозначения:
Р — родительские организмы
![]() — (алхимический знак Венеры — зеркало с ручкой) — материнская особь
— (алхимический знак Венеры — зеркало с ручкой) — материнская особь
![]() — (алхимический знак Марса — щит и копье) — отцовская особь
— (алхимический знак Марса — щит и копье) — отцовская особь
х — знак скрещивания
F1, F2, F3 и т. д. — гибриды первого, второго, третьего и последующих поколений;
Fа — потомство от анализирующего скрещивания.
Современные представления о гене и геноме
Гены, кодирующие первичную структуру белка, рибосомальной или транспортной РНК, называются структурными, а гены, обеспечивающие активацию или подавление считывания информации со структурных генов, — регуляторными.
Структура прокариотического гена. Структурный ген прокариот имеет сложное строение, поскольку в его состав входят регуляторные участки и кодирующие последовательности. К регуляторным участкам относятся промотор, оператор и терминатор (рис. 44). Промотором называют участок гена, к которому прикрепляется фермент РНК- полимераза, обеспечивающий синтез иРНК в процессе транскрипции. С оператором, располагающимся между промотором и структурной последовательностью, может связываться белок-репрессор, не позволяющий РНК-полимеразе начать считывание наследственной информации с кодирующей последовательности, и только его удаление позволяет начать транскрипцию. Структура репрессора закодирована обычно в регуляторном гене, находящемся в другом участке хромосомы. Считывание информации заканчивается на участке гена, который называется терминатором.

Рис. 44. Схема строения оперона:
1 — промотор; 2 — оператор; 3 — структурная последовательность; 4 — терминатор; 5 — РНК-полимераза; 6 — репрессор
Кодирующая последовательность структурного гена содержит информацию о последовательности аминокислот в соответствующем белке. Кодирующую последовательность у прокариот называют цистроном, а совокупность кодирующих и регуляторных участков гена прокариот — опероном. Структура оперона кишечной палочки и регуляция его активности были открыты Ф. Жакобом, Ж. Л. Моно и А. Львовым (1961 г.).
В целом прокариоты, к которым относится и кишечная палочка, имеют сравнительно небольшое количество генов, расположенных в единственной кольцевой хромосоме.
Структура эукариотического гена. В отличие от прокариот, гены эукариот не имеют оперонной структуры, поскольку не содержат оператора, и каждый структурный ген сопровождается только промотором и терминатором. Кроме того, в генах эукариот значащие участки (экзоны) чередуются с незначащими (интронами), которые полностью переписываются на иРНК, а затем вырезаются в процессе их созревания. Биологическая роль ин- тронов состоит в снижении вероятности мутаций в значащих участках. Регуляция генов эукариот намного сложнее, нежели описанная для прокариот.
Геном человека. В ходе международного проекта «Геном человека» (1988—2001 гг.) были осуществлена полная расшифровка последовательности нуклеотидов человеческой ДНК, обнаружение некоторых генов и картирование хромосом. Считается, что у человека 25—30 тыс. структурных генов, причем некоторые из них повторяются сотни и тысячи раз. При этом два разных человека на 99,9 % имеют сходные последовательности нуклеотидов, и лишь остающиеся 0,1 % определяют нашу индивидуальность. Вместе с тем расшифровка генома человека пока не дает прямого эффекта, поскольку мы получили своеобразную инструкцию по сборке такого сложного организма как человек, но не научились изготавливать его или хотя бы исправлять погрешности в нем.
Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание)
Основные закономерности независимого наследования признаков были открыты Г. Менделем, который достиг успеха, применив в своих исследованиях новый на тот момент гибридологический метод.
Для исследования Г. Мендель отобрал только семь признаков, имеющих альтернативные (контрастные) проявления. Уже в первых скрещиваниях он обратил внимание, что в потомстве первого поколения при скрещивании растений с желтыми и зелеными семенами все потомство имело желтые семена. Аналогичные результаты были получены и при исследовании других признаков (см. «Экспресс-помощник»).
Признаки, которые преобладали в первом поколении, Г. Мендель назвал доминантными. Те же из них, которые не проявлялись в первом поколении, получили название рецессивных.
Особи, которые давали расщепление в потомстве, получили название гетерозиготных, а особи, не дававшие расщепления, — гомозиготных.
Скрещивание, при котором исследуется проявление только одного признака, называется моногибридным. В таком случае прослеживаются закономерности наследования только двух вариантов одного признака, развитие которых обусловлено парой аллельных генов. Например, признак «окраска венчика цветка» у гороха имеет только два проявления — красная и белая. Все остальные признаки, свойственные данным организмам, во внимание не принимаются и не учитываются в расчетах.
Схема моногибридного скрещивания такова:

Скрестив два растения гороха, одно из которых имело желтые семена, а другое — зеленые, в первом поколении Г. Мендель получал растения исключительно с желтыми семенами, независимо от того, какое растение было выбрано в качестве материнского, а какое — отцовского. Такие же результаты были получены и в скрещиваниях по другим признакам, что дало Г. Менделю основания сформулировать закон единообразия гибридов первого поколения, который также называют первым законом Менделя и законом доминирования.
Первый закон Менделя:
При скрещивании гомозиготных родительских форм, отличающихся по одной паре альтернативных признаков, все гибриды первого поколения будут единообразны как по генотипу, так и по фенотипу.
А — желтые семена; а — зеленые семена.

При самоопылении (скрещивании) гибридов первого поколения оказалось, что 6 022 семени имеют желтую окраску, а 2 001 — зеленую, что примерно соответствует соотношению 3:1. Обнаруженная закономерность получила название закона расщепления, или второго закона Менделя.
Второй закон Менделя:
При скрещивании гетерозиготных гибридов первого поколения в потомстве будет наблюдаться преобладание одного из признаков в соотношении 3:1 по фенотипу (1:2:1 по генотипу).

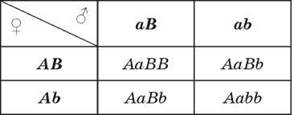
Однако по фенотипу особи далеко не всегда удается установить ее генотип, поскольку как гомозиготы по доминантному гену (АА), так и гетерозиготы (Аа) будут иметь в фенотипе проявление доминантного гена. Поэтому для организмов с перекрестным оплодотворением применяют анализирующее скрещивание — скрещивание, при котором организм с неизвестным генотипом скрещивается с гомозиготой по рецессивному гену для проверки генотипа. При этом гомозиготные особи по доминантному гену расщепления в потомстве не дают, тогда как в потомстве гетерозиготных наблюдается равное количество особей как с доминантным, так и с рецессивным признаками:

Основываясь на результатах собственных экспериментов, Г. Мендель предположил, что наследственные факторы при образовании гибридов не смешиваются, а сохраняются в неизменном виде. Поскольку связь между поколениями осуществляется через гаметы, то он допустил, что в процессе их образования в каждую из гамет попадает только один фактор из пары (то есть гаметы генетически чисты), а при оплодотворении пара восстанавливается. Эти предположения получили название правила чистоты гамет.
Правило чистоты гамет:
При гаметогенезе гены одной пары разделяются, то есть каждая гамета несет только одну аллель.
Однако организмы отличаются друг от друга по многим признакам, поэтому установить закономерности их наследования возможно только при анализе двух и более признаков в потомстве. Скрещивание, при котором рассматривается наследование и производится точный количественный учет потомства по двум парам признаков, называется дигибридным. Если же анализируется проявление большего числа наследственных признаков, то это уже полигибридное скрещивание.
Схема дигибридного скрещивания:

При большем разнообразии гамет определение генотипов потомков становится затруднительным, поэтому для анализа широко используется решетка Пеннета, в которую по горизонтали заносятся мужские гаметы, а по вертикали — женские. Генотипы потомков определяются сочетанием генов в столбцах и строках.
Для дигибридного скрещивания Г. Мендель выбрал два признака: окраску семян (желтую и зеленую) и их форму (гладкую и морщинистую). В первом поколении соблюдался закон единообразия гибридов первого поколения, а во втором поколении было 315 желтых гладких семян, 108 — зеленых гладких, 101 — желтое морщинистое и 32 зеленых морщинистых. Подсчет показал, что расщепление приближалось к 9:3:3:1, но по каждому из признаков сохранялось соотношение 3:1 (желтые — зеленые, гладкие — морщинистые). Эта закономерность получила название закона независимого расщепления признаков, или третьего закона Менделя.
Третий закон Менделя:
При скрещивании гомозиготных родительских форм, отличающихся по двум и более парам признаков, во втором поколении будет происходить независимое расщепление данных признаков в соотношении 3:1 (9:3:3:1 при дигибридном скрещивании).
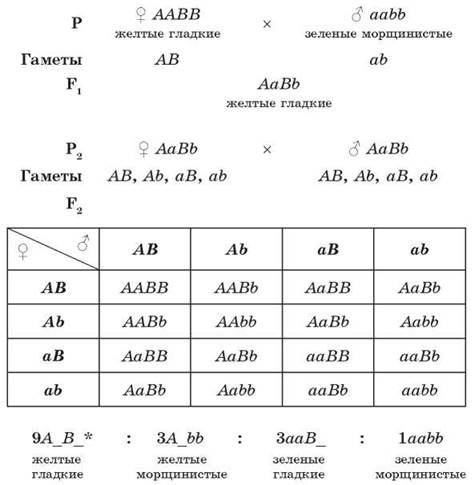
* На месте, обозначенном «_», может находиться как доминантный, так и рецессивный ген.
Третий закон Менделя применим только к случаям независимого наследования, когда гены расположены в разных парах гомологичных хромосом. В тех случаях, когда гены расположены в одной паре гомологичных хромосом, действительны закономерности сцепленного наследования. Закономерности независимого наследования признаков, установленные Г. Менделем, также часто нарушаются и при взаимодействии генов.
Закономерности наследственности, их цитологические основы
Согласно хромосомной теории наследственности каждая пара генов локализована в паре гомологичных хромосом, причем каждая из хромосом несет только по одному из этих факторов. Если представить, что гены являются точечными объектами на прямых — хромосомах, то схематически гомозиготные особи могут быть записаны как АllА или alla, тогда как гетерозиготная —Allа. При образовании гамет в процессе мейоза каждый из генов пары гетерозиготы окажется в одной из половых клеток (рис. 45).
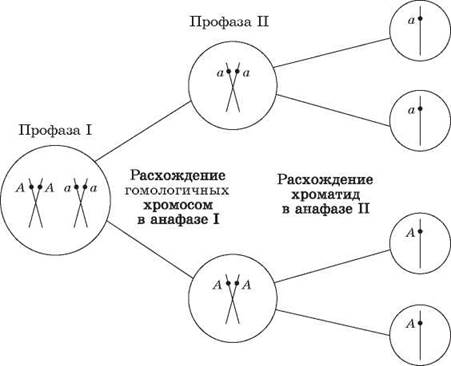
Рис. 45. Расхождение аллельных генов в процессе гаметогенеза
Например, если скрестить двух гетерозиготных особей, то при условии образования у каждой из них только пары гамет возможно получение всего лишь четырех дочерних организмов, три из которых будут нести хотя бы один доминантный ген А, и только один будет гомозиготен по рецессивному гену а, то есть закономерности наследственности носят статистический характер (рис. 46).
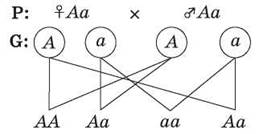
Рис. 46. Сочетания гамет в процессе оплодотворения
Если гены располагаются в разных хромосомах, то при образовании гамет распределение между ними аллелей из данной пары гомологичных хромосом происходит совершенно независимо от распределения аллелей из других пар.
Закономерности наследственности 146 148
Именно случайное расположение гомологичных хромосом на экваторе веретена в метафазе I мейоза и их последующее расхождение в анафазе I ведет к разнообразию рекомбинаций аллелей в гаметах (рис. 47).
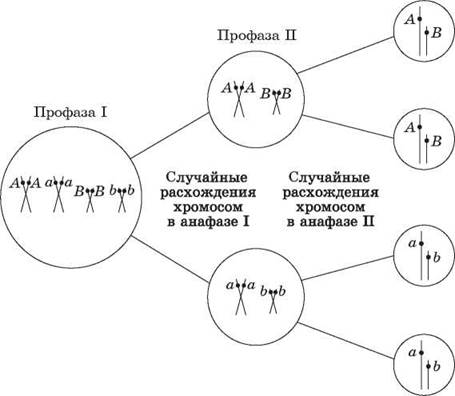
Рис. 47. Независимое расщепление каждой пары признаков
Число возможных сочетаний аллелей в мужских или женских гаметах можно определить по общей формуле 2n, где n — число хромосом, характерное для гаплоидного набора. У человека n- 23, а возможное число сочетаний составляет 223 = 8 388 608. Последующее объединение гамет при оплодотворении является также случайным, и поэтому в потомстве можно зафиксировать независимое расщепление по каждой паре признаков.
Однако число признаков у каждого организма во много раз больше числа его хромосом, которые можно различить под микроскопом, следовательно, каждая хромосома должна содержать множество факторов. Если представить себе, что у некоторой особи, гетерозиготной по двум парам генов, расположенных в гомологичных хромосомах, образуются гаметы, то следует учитывать не только вероятность образования гамет с исходными хромосомами, но и гамет, получивших измененные в результате кроссинговера в профазе I мейоза хромосомы. Следовательно, в потомстве возникнут новые сочетания признаков. Данные, полученные в экспериментах на дрозофиле, легли в основу хромосомной теории наследственности.
Закон Моргана: сцепленное наследование признаков, нарушение сцепления генов
Новый организм получает от родителей не россыпь генов, а целые хромосомы, при этом количество признаков и соответственно определяющих их генов гораздо больше, чем хромосом. В соответствии с хромосомной теорией наследственности, гены, расположенные в одной хромосоме, наследуются сцепленно. Вследствие этого при дигибридном скрещивании они не дают ожидаемого расщепления 9:3:3:1 и не подчиняются третьему закону Менделя. Можно было бы ожидать, что сцепление генов является полным, и при скрещивании гомозиготных по данным генам особей и во втором поколении дает исходные фенотипы в соотношении 3:1, а при анализирующем скрещивании гибридов первого поколения расщепление должно составлять 1:1.
Для проверки этого предположения американский генетик Т. Морган выбрал у дрозофилы пару генов, контролирующих окраску тела (серое — черное) и форму крыла (длинные — зачаточные), которые расположены в одной паре гомологичных хромосом. Серое тело и длинные крылья являются доминантными признаками. При скрещивании гомозиготной мухи с серым телом и длинными крыльями и гомозиготной мухи с черным телом и зачаточными крыльями во втором поколении действительно были получены в основном родительские фенотипы в соотношении, близком к 3:1, однако имелось и незначительное количество особей с новыми комбинациями этих признаков. Данные особи называются рекомбинантными.
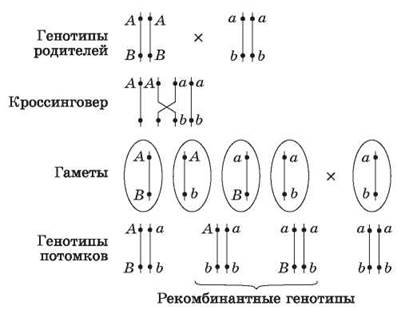
Рис. 48. Сцепленное наследование признаков и кроссинговер
Однако, проведя анализирующее скрещивание гибридов первого поколения с гомозиготами по рецессивным генам, Т. Морган обнаружил, что 41,5 % особей имели серое тело и длинные крылья, 41,5 % — черное тело и зачаточные крылья, 8,5 % — серое тело и зачаточные крылья, и 8,5 % — черное тело и зачаточные крылья. Он связал полученное расщепление с кроссинговером, происходящим в профазе I мейоза, и предложил считать единицей расстояния между генами в хромосоме 1 % кроссинговера, впоследствии названный в его честь морганидой.
Закономерности сцепленного наследования, установленные в ходе экспериментов на дрозофиле, получили название закона Моргана.
Закон Моргана:
Гены, локализованные в одной хромосоме, занимают определенное место, называемое локусом, и наследуются сцепленно, причем сила сцепления обратно пропорциональна расстоянию между генами.
Гены, расположенные в хромосоме непосредственно друг за другом (вероятность кроссинговера крайне мала), называются сцепленными полностью, а если между ними находится еще хотя бы один ген, то они сцеплены не полностью и их сцепление нарушается при кроссинговере в результате обмена участками гомологичных хромосом.
Хромосомная теория наследственности
Результаты экспериментов основоположника генетики Г. Менделя, английских генетиков У. Бэтсона, Р. Пеннета, Л. Донкастера и группы ученых, возглавляемой Т. Морганом, позволили к середине 20-х гг. XX в. сформулировать основные положения хромосомной теории наследственности, определить порядок расположения генов в хромосомах и расстояния между ними, то есть составить первые карты хромосом.
Основные положения хромосомной теории наследственности:
1. Гены расположены в хромосомах.
2. Гены одной хромосомы наследуются совместно, или сцепленно, и называются группой сцепления.
3. Число групп сцепления численно равно гаплоидному набору хромосом.
4. В каждой хромосоме сосредоточены определенные гены, отвечающие за целый ряд признаков.
5. Каждый ген занимает в хромосоме строго определенное место — локус.
6. Гены в хромосомах расположены линейно.
7. Нарушение сцепления генов происходит только в результате кроссинговера.
8. Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними.
9. Независимое наследование характерно только для генов негомологичных хромосом.
Генетика пола
Пол — это совокупность морфологических и физиологических особенностей организма, обеспечивающих половое размножение, сущность которого сводится к оплодотворению, то есть слиянию мужских и женских половых клеток в зиготу, из которой развивается новый организм.
Признаки, по которым один пол отличается от другого, делят на первичные и вторичные. К первичным половым признакам относятся половые органы, а все остальные — ко вторичным.
У человека вторичными половыми признаками являются тип телосложения, тембр голоса, преобладание мышечной или жировой ткани, наличие оволосения на лице, кадыка, молочных желез. Так, у женщин таз обычно шире плеч, преобладает жировая ткань, выражены молочные железы, голос высокий. Мужчины же отличаются от них более широкими плечами, преобладанием мышечной ткани, наличием оволосения на лице и кадыка, а также низким голосом.
В кариотипе мужчин есть две непарные хромосомы. По форме эти хромосомы в начале деления напоминают латинские буквы X и Y, и поэтому были названы X- и Y-хромосомами. Сперматозоиды мужчины могут нести одну из этих хромосом и определять пол будущего ребенка. В связи с этим хромосомы человека и многих других организмов делят на две группы: аутосомы и гетерохромосомы, или половые хромосомы.
В тех случаях, когда пол несет одинаковые половые хромосомы, например, XX, говорят, что он гомозиготен, или гомогаметен (образует одинаковые гаметы). Другой же пол, имеющий разные половые хромосомы (XY), называется гемизиготным (не имеющим полного эквивалента аллельных генов), или гетерогаметным. У человека, большинства млекопитающих, мушки дрозофилы и других организмов гомогаметен женский пол (XX), а мужской — гетерогаметен (XY), тогда как у птиц гомогаметен мужской пол (ZZ, или XX), а женский — гетерогаметен (ZW, илиXY).
Кариотип мужчины записывается как ![]() 46, XY, а кариотип женщины — как
46, XY, а кариотип женщины — как ![]() 46, XX.
46, XX.
Поскольку некоторые гены находятся в половых хромосомах, неодинаковых у представителей противоположных полов, характер наследования признаков, кодируемых данными генами, отличается от общего. Такой тип наследования называется крис-кросс наследованием, поскольку мужчины наследуют признаки матери, а женщины — отца. Признаки, определяемые генами, которые находятся в половых хромосомах, называются сцепленными с полом. Примерами признаков, сцепленных с полом, являются рецессивные признаки гемофилии и дальтонизма, которые в основном проявляются у мужчин, так как в Y-хромосоме нет аллельных генов. Женщины болеют такими болезнями только в том случае, если и от отца, и от матери они получили такие признаки.
Например, если мать была гетерозиготным носителем гемофилии, то у половины ее сыновей свертываемость крови будет нарушена:
XH — нормальное свертывание крови;
Xh — несвертываемость крови (гемофилия).

Признаки, закодированные в генах У-хромо- сомы, передаются сугубо по мужской линии и называются голандрическими (наличие перепонки между пальцами ног, повышенное оволосение края ушной раковины).
Генотип как целостная система. Взаимодействие генов
Генотип не является механической суммой генов, поскольку возможность проявления гена и форма его проявления зависят от условий среды. В данном случае под средой понимается не только окружающая среда, но и генотипическая среда — другие гены.
Проявление качественных признаков редко зависит от условий окружающей среды, хотя, если у горностаевого кролика выбрить участок тела с белой шерстью и прикладывать к нему пузырь со льдом, то со временем на этом месте вырастет черная шерсть.
Развитие количественных признаков намного сильнее зависит от условий окружающей среды, например, если современные сорта пшеницы возделывать без применения минеральных удобрений, то ее урожайность будет существенно отличаться от генетически запрограммированных 100 и более центнеров с гектара.
Таким образом, в генотипе записаны лишь «способности» организма, однако проявляются они только во взаимодействии с условиями окружающей среды.
Кроме того, гены взаимодействуют друг с другом и, оказавшись в одном генотипе, могут сильно влиять на проявление действия соседних генов.
В связи с этим известный российский генетик М. Е. Лобашев определил генотип как систему взаимодействующих генов. Сложилась эта целостная система в процессе эволюции органического мира, при этом выживали лишь те организмы, у которых взаимодействие генов давало наиболее благоприятную реакцию в онтогенезе.
Ко взаимодействию аллельных генов относят полное и неполное доминирование, кодоминирование и сверхдоминирование. Полным доминированием считают все случаи взаимодействия аллельных генов, при которых в гетерозиготе наблюдается проявление исключительно доминантного признака, как, например, окраска и форма семени у гороха.
Неполное доминирование — это тип взаимодействия аллельных генов, при котором проявление рецессивного аллеля в большей или меньшей степени ослабляет проявление доминантного, как в случае окраски венчика ночной красавицы (белая + красная = розовая) и шерсти у крупного рогатого скота.
Кодоминированием называют такой тип взаимодействия аллельных генов, при котором оба аллеля проявляются, не ослабляя эффектов друг друга. Типичным примером кодоминирования является наследование групп крови по системе AB0 (табл. 2). IV (АВ) группа крови у человека (генотип — IAIB).
Как видно из таблицы, I, II и III группы крови наследуются по типу полного доминирования, тогда как IV (АВ) группа (генотип — IAIB) является случаем кодоминирования.
Сверхдоминирование — это явление, при котором в гетерозиготном состоянии доминантный признак проявляется намного сильнее, чем в гомозиготном; сверхдоминирование часто используется в селекции и считается причиной гетерозиса — явления гибридной силы.
Особым случаем взаимодействия аллельных генов можно считать так называемые летальные гены, которые в гомозиготном состоянии приводят к гибели организма чаще всего в эмбриональном периоде. Причиной гибели потомства является плейотропное действие генов серой окраски шерсти у каракулевых овец, платиновой окраски у лис и отсутствие чешуи у зеркальных карпов. При скрещивании двух гетерозиготных по этим генам особей расщепление по исследуемому признаку в потомстве будет равняться 2:1 вследствие гибели 1/4 потомства.
Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия. Комплементарность — это тип взаимодействия неаллельных генов, при котором для проявления определенного состояния признака необходимо присутствие как минимум двух доминантных аллелей разных пар, например, у тыквы при скрещивании растений со сферическими (ААbb) и длинными (aaBB) плодами в первом поколении появляются растения с дисковидными плодами (AaBb).
К эпистазу относят такие явления взаимодействия неаллельных генов, при которых один неаллельный ген подавляет развитие признака другого, например, у кур окраска оперения определяется одним доминантным геном, тогда как другой доминантный ген подавляет развитие окраски, в результате чего большинство кур имеют белое оперение.
Полимерией называют явление, при котором неаллельные гены оказывают одинаковое влияние на развитие признака. Таким образом чаще всего кодируются количественные признаки, например, цвет кожи человека определяется как минимум четырьмя парами неаллельных генов — чем больше доминантных аллелей в генотипе, тем темнее кожа.
Генетика человека
Для человека как биологического вида в полной мере справедливы генетические закономерности наследственности и изменчивости, установленные для растений и животных. Вместе с тем генетика человека, изучающая закономерности наследственности и изменчивости у человека на всех уровнях его организации и существования, занимает особое место среди других разделов генетики.
Биосоциальная природа человека накладывает значительный отпечаток на исследования в области его генетики вследствие позднего полового созревания и больших временных разрывов между поколениями, малочисленности потомства, невозможности направленных скрещиваний для генетического анализа, отсутствия чистых линий, недостаточной точности регистрации наследственных признаков и небольших родословных, невозможности создания одинаковых и строго контролируемых условий для развития потомков от разных браков, сравнительно большого числа плохо различающихся хромосом и невозможности экспериментального получения мутаций.
Методы, применяемые в генетике человека, принципиально не отличаются от общепринятых для других объектов — это: генеалогический, близнецовый, цитогенетический, дерматоглифический, молекулярно-биологический и популяционно-статистический методы, метод гибридизации соматических клеток и метод моделирования. Их использование в генетике человека учитывает специфику человека как генетического объекта.
ЭТО ИНТЕРЕСНО. Генетика человека занимается исследованием наследственных болезней, которых описано уже более 4 тыс.
Генеалогический метод — это метод составления родословных, с помощью которых определяют характер наследования изучаемых признаков, в том числе наследственных болезней, и прогнозируют рождение потомков с соответствующими признаками. Он позволил выявить наследственную природу таких заболеваний, как гемофилия и дальтонизм, еще до открытия основных закономерностей наследственности. При составлении родословных ведут записи о каждом из членов семьи и учитывают степень родства между ними. Далее на основании полученных данных с помощью специальной символики строится родословное древо.
Генеалогический метод можно использовать на одной семье, если есть сведения о достаточном количестве прямых родственников человека, родословная которого — пробанда — составляется по отцовской и материнской линиям, в противном случае собирают сведения о нескольких семьях, в которых проявляется данный признак. Генеалогический метод позволяет установить не только наследуемость признака, но и характер наследования: доминантный или рецессивный, аутосомный или сцепленный с полом и т. д.
При составлении родословных принято использовать условные обозначения (рис. 49).
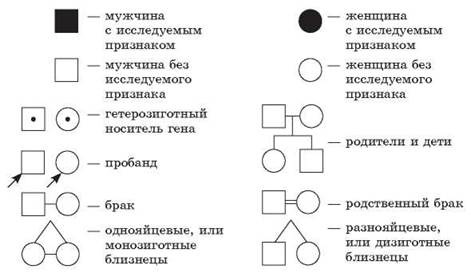
Рис. 49. Условные обозначения, используемые при составлении родословных
Близнецовый метод помогает определить вклад наследственности и влияние условий окружающей среды на проявление признака на основе анализа совпадения этих признаков у однояйцевых и разнояйцевых близнецов. Так, у большинства однояйцевых близнецов совпадают группы крови, цвет глаз и волос, а также целый ряд других признаков, тогда как корью болеют одновременно оба типа близнецов.
Дерматоглифический метод основан на исследовании индивидуальных особенностей кожных рисунков пальцев рук (дактилоскопия), ладоней и ступней ног. На основе этих особенностей он зачастую позволяет своевременно выявить наследственные заболевания, в частности хромосомные аномалии, такие как синдром Дауна, Шерешевского—Тернера и др.
Закономерности изменчивости
Давно было подмечено, что даже однояйцевые близнецы-клоны, выросшие в разных условиях, приобретают целый ряд различий, обусловленных, прежде всего, условиями среды. Это свойство организмов приобретать новые признаки в течение жизни называется изменчивостью. Ее изучение является одной из основных задач генетики, так как выявление причин и пределов изменчивости позволяет установить соотношение наследственных задатков и приобретенных отклонений в проявлении признака, создать оптимальные условия для реализации генетически заложенного потенциала породами животных и сортами растений, а также предотвратить проявление нежелательных признаков.
Изменчивость делят на наследственную, или генотипическую, и ненаследственную, или фенотипическую.
Наследственная изменчивость закрепляется в генотипе и передается в ряду поколений. К наследственной изменчивости относят комбинативную и мутационную изменчивости. Она является основой разнообразия живых организмов и эволюции в целом.
Ненаследственная, или групповая, изменчивость не связана с изменениями генотипа и не передается в ряду поколений. К ней относится прежде всего модификационная изменчивость.
Ненаследственная (модификационная) изменчивость
Фенотипические изменения организма, не связанные с изменениями наследственного аппарата, называют модификациями.
Модификации являются результатом взаимодействия генотипа и окружающей среды. Примерами модификационной изменчивости является разнолистность у растений, окраска крыльев у бабочки пестрокрыльницы изменчивой, загар у человека и т. д. Так, у стрелолиста надводные листья имеют традиционную стреловидную форму, плавающие — округлую, а подводные — удлиненную.
Модификации имеют приспособительное значение, поскольку у стрелолиста лентовидный лист менее подвержен риску повреждения течением, а загар защищает организм человека от губительного действия ультрафиолетовых лучей. Скорее всего модификации возникли в процессе эволюции каждого вида как адаптации к наиболее часто встречающимся изменениям факторов окружающей среды. При попадании организма в непривычные условия могут возникать модификации, лишенные биологического смысла, например, при затенении нижней части стебля картофеля на нем начинают образовываться надземные клубни.
При всей пластичности признаков живых организмов даже модификационная изменчивость имеет определенные пределы, обусловленные возможностями генотипа.
Норма реакции — это диапазон фенотипических проявлений одного и того же генотипа в разных условиях среды.
Норма реакции, по-видимому, сформировалась в процессе эволюции, поскольку в конкретных условиях среды выживали только те организмы, фенотипические изменения которых способствовали их существованию, то есть норма реакции имеет приспособительный характер.
Исследовав конкретный признак у многих особей, можно составить вариационный ряд — последовательность количественных показателей состояния признака, или вариант, расположенных в порядке их возрастания или убывания. Длина данной последовательности будет зависеть как от пластичности генотипа, так и от изменчивости условий среды. Распределение отдельных вариантов в популяции носит статистический характер, поскольку наиболее часто встречается среднее значение признака, тогда как его крайние проявления достаточно редки, что обусловлено более редким воздействием экстремальных значений факторов среды. Например, если измерить листья одного из видов эвкалипта, то на 100 листьев довольно редко встречаются не достигающие в длину 14 см или превышающие 20 см, тогда как средние размеры колеблются в пределах 16—18 см. На основании измерений или взвешиваний строятся графические отображения изменчивости признака, отражающие норму реакции и частоту встречаемости отдельных вариант — вариационные кривые (рис. 50).
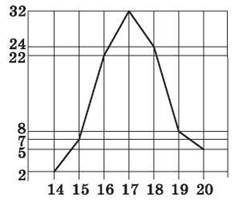
Рис. 50. Вариационная кривая количества колосков в колосе пшеницы
Наследственная изменчивость: мутационная, комбинативная
Наследственная изменчивость закрепляется в генотипе и передается потомкам. Она является основой разнообразия живых организмов и эволюции в целом. К наследственной изменчивости относят, прежде всего, комбинативную и мутационную изменчивости.
Комбинативная изменчивость — форма наследственной изменчивости, обусловленная сочетанием генов, полученных от обоих родителей в процессе оплодотворения.
Она возникает как результат рекомбинации генов при кроссинговере, независимого расхождения хромосом в анафазе I мейоза и случайной встречи гамет при оплодотворении. Все генотипическое и фенотипическое разнообразие особей любой систематической группы является результатом комбинативной изменчивости, поскольку только в этом процессе возникают уникальные сочетания генов и устанавливаются новые взаимодействия между ними, что в конечном итоге и определяет появление новых признаков и свойств организмов.
Комбинативная изменчивость может наблюдаться и у организмов, размножающихся бесполым способом, например, у бактерий при переносе информации бактериофагами от одной бактерии к другой в процессе заражения.
Мутационная изменчивость — это форма изменчивости, связанная с возникновением и проявлением мутаций.
Мутация — это внезапно возникающее естественное или вызванное искусственно изменение генетического материала, приводящее к изменению тех или иных наследственных признаков организма.
Существует несколько классификаций мутаций. По месту возникновения их делят на соматические и генеративные. Наиболее опасными являются генеративные мутации, так как они не отражаются на жизнедеятельности родительских особей, проявляясь только у потомков.
По влиянию на жизнеспособность организмов выделяют летальные, полулетальные и нейтральные мутации. Летальные мутации вызывают гибель организма чаще всего в эмбриональном периоде развития, как, например, у некоторых каракулевых овец. Полулетальные, или сублетальные, мутации снижают жизнеспособность особей (серповидно-клеточная анемия у человека), тогда как нейтральные мутации не влияют на их жизнеспособность.
В зависимости от того, насколько большой участок наследственной информации затрагивает мутация, выделяют генные, хромосомные и геномные мутации. Генные, или точечные, мутации — это мутации, связанные с перестановками нуклеотидов, их выпадением и вставками в пределах одного гена. Многие генные мутации приводят к тяжелым генным заболеваниям, таким как гемофилия, дальтонизм, серповидно-клеточная анемия и др.
Хромосомные мутации вызваны изменениями более крупных участков ДНК, затрагивающих уже несколько генов. Примером болезни, вызванной такой мутацией, является синдром «кошачьего крика».
Геномными называют мутации, связанные с изменением числа хромосом, то есть их нехваткой или избытком. Изменение количества хромосом на число, кратное гаплоидному набору хромосом, называется полиплоидией. Полиплоидизация широко используется при выведении новых сортов растений, так как они обычно обладают большей продуктивностью, чем диплоидные растения.
Явлением, обратным полиплоидизации, является гаплоидизация. Несмотря на то что гаплоидные формы обычно имеют меньшие размеры и сниженную продуктивность, их используют в селекции для выявления рецессивных аллелей и получения форм, гомозиготных по всем генам.
Изменение числа хромосом на некратное гаплоидному набору называется анеуплоидией. У животных и человека анеуплоидии вызывают тяжелые расстройства состояния здоровья и часто смертельно опасны. Примерами анеуплоидий у человека являются синдромы Дауна, Шерешевского—Тернера, Клайнфельтера.
Мутации возникают спонтанно (под действием внутренних факторов) или под действием различных факторов, которые называют мутагенами. Мутагены делят на физические, химические и биологические. В качестве физических мутагенов могут выступать ионизирующая радиация, ультрафиолетовое излучение, повышенные температуры и др. Химическими мутагенами являются не только различные органические растворители, нефтепродукты, соли тяжелых металлов, многие лекарственные препараты, но и алкоголь, никотин, наркотические вещества. К биологическим мутагенам относят, прежде всего, вирусы, способные встраиваться в наследственный аппарат клетки, повреждая его, а также переносить фрагменты другим организмам, как это происходит у бактерий с помощью бактериофагов.
Значение изменчивости в жизни организмов и в эволюции
Изменчивость играет важную роль в процессах как индивидуального развития организмов, так и в их историческом развитии. Особое место в этом процессе занимают мутации, поскольку они могут оказаться как полезными, так и вредными, в том числе и летальными. Так, у человека значительная часть беременностей заканчивается самопроизвольным выкидышем на ранних сроках вследствие наличия у зародыша геномных и хромосомных аномалий. Хромосомные мутации могут также привести к возникновению и закреплению новых благоприятных признаков, являющихся результатом взаимодействия генов, которые оказались рядом.
Генные мутации обеспечивают полиморфмизм аллелей в определенных локусах, что увеличивает гетерозиготность популяции, делает более разнообразным ее генофонд и ведет к усилению внутрипопуляционной изменчивости. При этом доминантные мутации сравнительно редко закрепляются в потомстве, поскольку особи с такими признаками непосредственно подвергаются естественному отбору, тогда как рецессивные мутации могут длительное время пребывать в популяции в скрытом состоянии и становятся объектом отбора только в том случае, когда их концентрация в популяции достигнет определенного значения и начнут появляться особи с рецессивными признаками.
Таким образом, мутационная изменчивость обеспечивает резерв выживаемости данного вида в меняющихся условиях среды.
Новые сочетания генов, которые создаются в результате кроссинговера, независимого расхождения хромосом и случайности встречи гамет (комбинативная изменчивость), с одной стороны, обеспечивают многообразие генотипов и фенотипов в популяциях, а с другой — являются преходящими, то есть так же легко создаются, как и разрушаются. Это явление объясняет появление у выдающихся родителей посредственных потомков.
Модификационная изменчивость не наследуется, однако она является своеобразной проверкой правильности выбранной стратегии адаптации к определенным условиям среды, и впоследствии возможно появление мутаций, которые бы закрепляли генетически лучшие варианты модификаций.
Селекция
Селекция — это наука о выведении сортов растений, пород животных и штаммов микроорганизмов с ценными для человека свойствами. Под селекцией подразумевают также и сам процесс их выведения.
Первым этапом селекции было одомашнивание (доместикация), в процессе которого шел отбор по поведению животных и способности размножаться под контролем человека. Оно позволило сохранить огромное разнообразие признаков, в том числе неблагоприятных для вида.
Выдающийся русский генетик и селекционер Н. И. Вавилов в ходе многочисленных экспедиций изучил мировые растительные ресурсы и установил, что наибольшее разнообразие форм определенного вида характерно для тех районов, где этот вид был введен в культуру. В соответствии с этим он определил семь центров происхождения культурных растений (табл. 3).
Дальнейшие исследования позволили выделить уже 12 центров происхождения культурных растений, тесно связанных с центрами одомашнивания животных.
На основании изучения признаков культурных растений и близких к ним диких видов Н. И. Вавилов в 1920 году сформулировал закон гомологических рядов в наследственной изменчивости:
Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство.
Генетической основой данного закона является то, что степень исторического родства прямо пропорциональна количеству их общих генов, вследствие чего и мутации этих генов могут быть сходными. В фенотипе это проявляется одинаковым характером изменчивости многих признаков у близких видов, родов и других таксонов.
Закон гомологических рядов наследственной изменчивости организмов объясняет направленность исторического развития родственных групп организмов. Опираясь на него и изучив наследственную изменчивость близких видов, в селекции планируют работу по созданию новых сортов растений и пород животных с определенным набором наследственных признаков. В систематике организмов этот закон позволяет предвидеть существование неизвестных науке систематических групп (видов, родов и т. д.) с подобными сочетаниями признаков, выявленных в близкородственных группах.
Основные методы селекции — гибридизация и искусственный отбор.
Гибридизация — это процесс образования или получения гибридов, в основе которого лежит объединение генетического материала разных клеток в одной клетке.
Для достижения результата в процессе гибридизации особое внимание уделяется подбору родительских пар. В селекции растений подбор ведется по определенным признакам с учетом генетической и географической удаленности; в селекции животных — только по хозяйственно ценным признакам, которые определяют по экстерьеру, родословной и потомству.
Выделяют родственную и неродственную гибридизации. Родственное скрещивание, или инбридинг, приводит к появлению чистых линий, но при этом снижается жизнеспособность потомства вследствие перехода различных летальных и полулетальных генов в гомозиготное состояние.
Неродственное скрещивание, или аутбридинг, бывает внутривидовым и межвидовым (в т. ч. отдаленная гибридизация). Аутбридинг в первом поколении дает эффект гетерозиса.
Гетерозис — явление повышения жизнеспособности и продуктивности у гибридов первого поколения по сравнению с исходными родительскими формами.
Данное явление объясняется благоприятным сочетанием родительских генов, а также переходом сублетальных и летальных аллелей в гетерозиготное состояние. Во втором и последующих поколениях эффект гетерозиса ослабевает вследствие расщепления генов и гомоготизации. Эффект гетерозиса широко применяется в сельском хозяйстве, так как он позволяет существенно повысить урожайность растений (кукурузы, огурцов, томатов) и продуктивность животных (яйценоскость гибридов леггорнов, скорость роста и улучшение качества мяса бройлеров).
Искусственный отбор — процесс создания новых пород животных и сортов культурных растений путем систематического сохранения и размножения особей с определенными, ценными для человека признаками и свойствами в ряду поколений.
Выделяют две формы искусственного отбора: бессознательный, ведущийся без определенного плана, и методический, производимый с определенной целью. Примером искусственного отбора являются породы домашних голубей, выведенные от дикого скалистого голубя. Также он применяется в форме массового и индивидуального отбора. Массовый отбор является эффективным при высокой наследуемости признака. В основном он используется в селекции растений и микроорганизмов. При индивидуальном отборе учитываются не только показатели продуктивности или иные качества организма, но и наследование данного признака в ряду поколений. В комбинации с инбридингом он позволяет получить чистые линии. Индивидуальный отбор характерен для селекции животных и самоопыляющихся растений.
Теорию искусственного отбора создал великий английский ученый Ч. Дарвин.