Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Резюме - Раздел 5 - Биогеография континентальных водоёмов
Биоту каждого биогеографического региона, как обычно, определяет история: исходная биота и возможность элементам других биот попасть на акваторию данной. В целом биогеографическое районирование пресных вод сходно с биогеографическим районированием суши. Вместе с тем имеются и существенные отличия.
Основой биогеографии суши является растительность, в биогеографии же континентальных водоёмов данные по растительности не имеют почти никакого значения.
Широтная зональность имеет свою специфику, связанную как с древностью многих пресноводных таксонов, так и их малой по сравнению с наземными таксонами, расселительной способностью. В континентальных водоёмах столь ясно выраженное увеличение видового разнообразия в низких широтах есть только среди некоторых первично пресноводных таксонов (Рис. 4.35, Табл. 4.9). Районы высоких широт, биота которых не была существенно обеднена в периоды оледенений обладают видовым разнообразием, не уступающим таковому тропиков, наиболее ярко выражено это в бассейне Миссисипи (Рис. 4.36, Рис. Ц8).
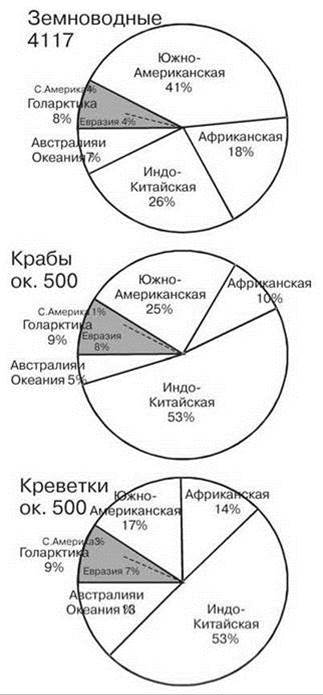
Рис. 4.35. Число видов амфибий и крабов в разных биогеографических регионах.
Земноводные Amphibia (только чисто водные виды) по Vences, Kohler (2008), крабы Brachyura по Yeo et al. (2008), креветки Caridea по Grave et al. (2008). Цифры при названии таксона — число видов.
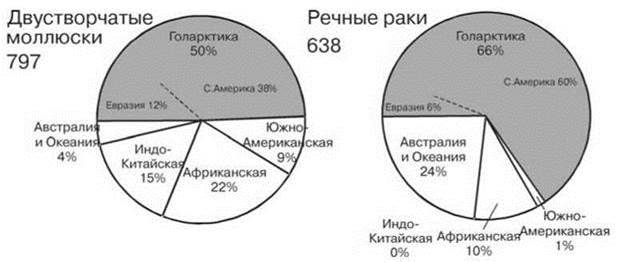
Рис. 4.36. Число видов двух отрядов ракообразных в разных биогеографических регионах.
Равноногие раки Isopoda по Wilson (2008), бокоплавы Amphipoda по Vainola et al. (2008). Цифры при названии таксона — число видов.
Табл. 4.9. Общее число видов основных таксонов пресноводных животных в разных районах (по Balian et al., 2008).
|
PA |
NA |
AT |
NT |
OL |
AU |
PAC |
ANT |
Мир |
|
|
Annelida |
870 |
350 |
186 |
338 |
242 |
210 |
10 |
10 |
1,761 |
|
Mollusca |
1,848 |
936 |
483 |
759 |
756 |
557 |
171 |
0 |
4,998 |
|
Crustacea |
4,499 |
1,755 |
1,536 |
1,925 |
1,968 |
1,225 |
125 |
33 |
11,990 |
|
Arachnida |
1,703 |
1,069 |
801 |
1,330 |
569 |
708 |
5 |
2 |
6,149 |
|
Insecta* |
15,190 |
9,410 |
8,594 |
14,428 |
13,912 |
7,510 |
577 |
14 |
75,874 |
|
Vertebrata** |
2,193 |
1,831 |
3,995 |
6,041 |
3,674 |
694 |
8 |
1 |
18,235 |
|
прочие |
4,013 |
1,721 |
1,194 |
1,365 |
1,239 |
956 |
184 |
112 |
6,523 |
|
Итого |
30,316 |
17,072 |
16,789 26,186 22,360 |
11,860 |
1,080 |
174 |
125,530 |
||
Биогеографические области по схеме наземной биоты (Cox, 2001): PA — Палеарктика, NA — Неарктика, AT — Афротропическая; OL — Ориентальная; AU — Австралазийская; PAC — Пацифическая; ANT — Антарктическая.
* Данные по некоторым семействам двукрылых неполны.
** Включены только строго пресноводные рыбы (кроме того есть около 2,300 солоноватоводных).
Высокое таксономическое разнообразие тропических регионов, в первую очередь, обеспечивают первично и вторично пресноводные таксоны. Многие древние первично пресноводные таксоны часто ограничены приэкваториальной зоной, хотя такие таксоны есть и в умеренных районах. Роль периферических таксонов возрастает только в районах, первично- и вторично-пресноводная фауна которых объединена: Центральная Америка и острова Напротив, фауна умеренных районов Северного полушария, в первую очередь облигатно пресноводных, значительно менее разнообразна, нежели тропическая, а многие тропические таксоны полностью отсутствуют. В этом континентальные водоёмы сходны с сушей, бенталью и пелагиалью. Но некоторые первично и облагитно пресноводные таксоны наоборот, в умеренных районах гораздо более разнообразны, чем в тропиках. Подобные явления известны и в других частных биогеографиях, но там они обычно свойственны таксонам низкого ранга или олиготипичным, а не богатым видами таксонам высокого ранга, как это наблюдается в континентальных водоёмах. Таковы бокоплавы и изоподы, особенно первично пресноводные (Asellota и Phreatoicidea) (Рис. 4.37). Высокое разнообразие изопод в Южной Америке наполовину обусловлено высоким разнообразием эктопаразитов рыб (Cymothoidae, 40 видов), в Африке 77% видов обитают в Южноафриканской провинции, а в тропической Азии 2/3 — на Индостане, это те же таксоны, что и в Южной Африке, т. е. они, вероятно, попали в Азию с Индостаном. Из десятиногих ракообразных: речные раки обитают только в умеренных широтах (кроме Австралии, причины этого рассмотрены выше), тогда как крабы (Рис. 4.35) и креветки приурочены к тропикам (за исключением Potaminae, приуроченных к восточному Тетису) (Darren, Peter, 2004).
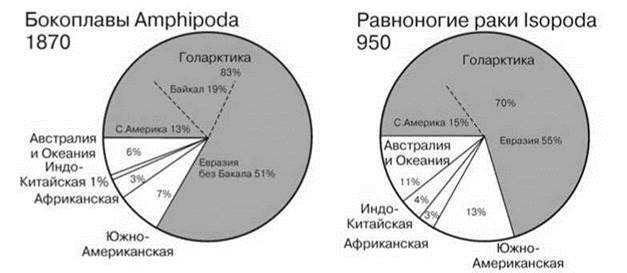
Рис. 4.37. Число видов двух отрядов ракообразных в разных биогеографических регионах.
Равноногие раки Isopoda по Wilson (2008), бокоплавы Amphipoda по Vainola et al. (2008). Цифры при названии таксона — число видов.
Любопытно также сравнительно равномерное распределение амфибийных виды, и, в частности, насекомых по областям (Рис. 4.38—4.40). Разнообразие одних групп в Голарктике значительно выше, чем в других областях, особенно веснянок. Разнообразные как в постоянных, так и во временных водоёмах жуки, ручейники и подёнки разнообразны и в Голарктике (29—47% мировой фауны). Стрекозы, бабочки и клопы дают практически идентичную картину: они сравнительно бедны в Голарктике (15—18% мировой фауны). Можно даже утверждать, что водные насекомые, в отличие от наземных, разнообразнее в умеренных широтах. Возможно, это связано с тем, что амфибийные виды — большей частью ценофобы, приуроченные к эфемерным водоёмам и водоёмам с высокоизменчивыми условиями. В стабильных условиях ЦС крупных озёр безраздельно доминируют ценофильные виды, роль насекомых там незначительна. В условиях умеренного климата к высокоизменчивым и/или эфемерным водоёмам относится большая часть мелких водоёмов (лужи, родники, ручьи, речушки). В низких широтах из-за менее изменчивого климата биота подобных водоёмов может включать больше ценофильных видов.
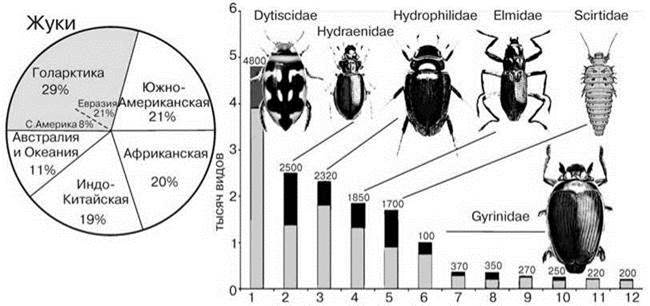
Рис. 4.38. Число видо жуков в разных биогеографических регионах и ожидаемое число видов семейств водных жуков в мировой фауне.
1 — Dytiscidae, 2 — Hydraenidae, 3 — Hydrophilidae, 4 — Elmidae, 5 — Scirtidae, 6 — Gyrinidae, 7 — Psephenidae, 8 — Dryopidae, 9 — Noteridae, 10 — Hydrochidae, 11 — Haliplidae, 12 — Helophoridae (по: Jаch, Balke, 2008). Цифры при названии таксона — число видов. По мнению Yee (2014) Dytiscidae только 4200 видов.
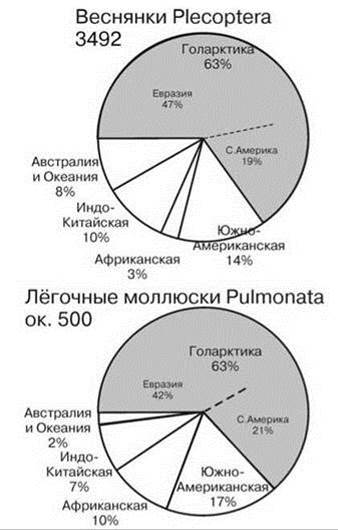
Рис. 4.39. Число видов веснянок и лёгочных моллюсков в разных биогеографических регионах. По Fochetti, Figueroa (2008) и Strong et al. (2008).
Цифры при названии таксона — число видов.
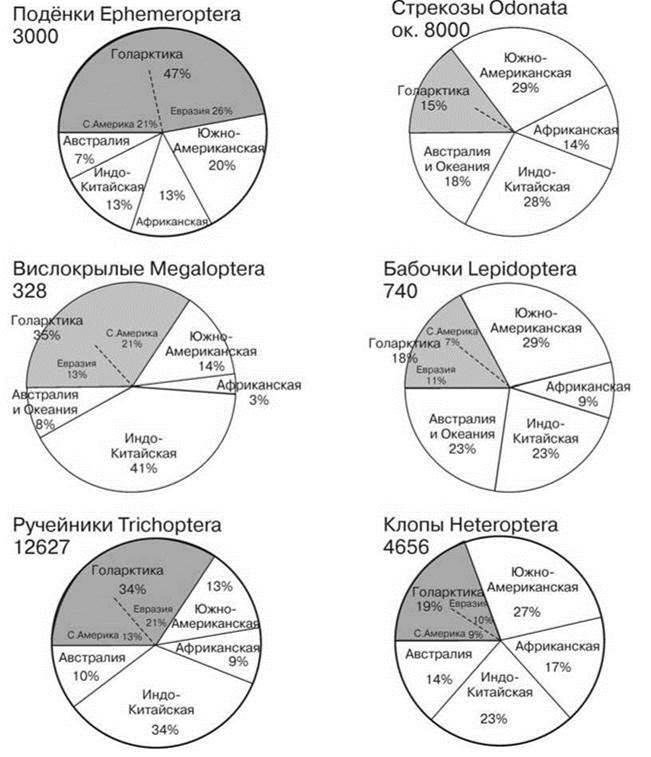
Рис. 4.40. Число видов разных отрядов насекомых с личинками и/или imago обитающими в воде в разных биогеографических регионах.
По: подёнки — Barber-James (2008), вислокрылки — Cover, Resh (2007), ручейники — Moor, Ivanov (2008), стрекозы — Kalkman et al. (2008), бабочки — Mey, Speidel (2008), клопы — Polhemus, Polhemus (2008). Цифры при названии таксона — число видов.
Распределение отрядов насекомых по постоянным и временным водоёмам показано на Рис. 4.41. На Рис. 4.42 показано распределение отрядов насекомых по водотокам разного типа.
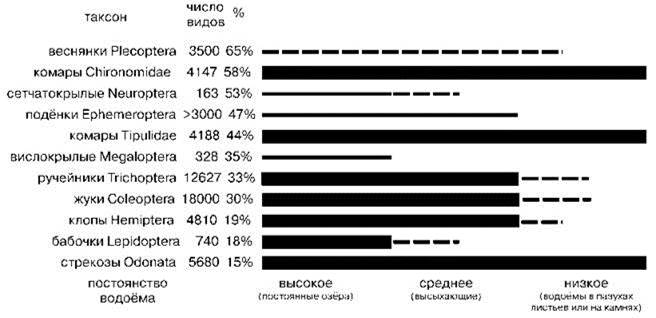
Рис. 4.41. Относительное число видов разных отрядов насекомых, обитающих в водоёмах разного типа (в скобках приведены примеры водоёмов).
Толстая линия означает, что в водоёмах данного типа виды таксона многочисленны, тонкая — относительно многочисленны, пунктирная — имеются, но немногочисленны. Таксоны упорядочены по доле голарктических видов в мировой фауне % справа от общего числа видов. По Williams (2006), число видов по Hydrobiologia (2008).
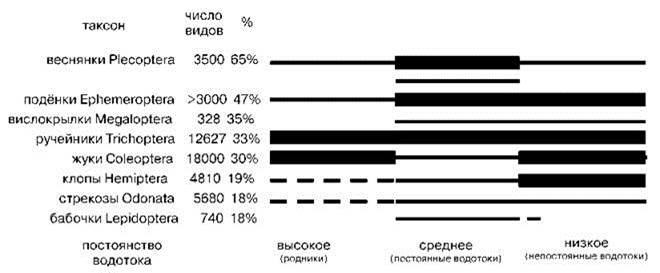
Рис. 4.42. Относительное число видов разных отрядов насекомых, обитающих в водотоках разного типа (в скобках приведены примеры водоёмов).
Толстая линия означает, что в водоёмах данного типа виды таксона многочисленны, тонкая — относительно многочисленны, пунктирная — имеются, но немногочисленны. По Williams(2006), число видов по Hydrobiologia (2008).
Сходно распределены Gastropoda. Логично допустить, что дышащие воздухом гастроподы более приурочены к нарушенным местообитаниям, нежели дышащие жабрами, т.е. среди первых доля ценофобов, вероятно, существенно выше. Как отмечено выше в оз. Танганьика дышащие воздухом гастроподы (прудовики Lymnaeidae, катушки Planorbidae, ампулярии Ampullariidae = Pilidae) обитают за пределами специфических ЦС озера, а эндемы имеются только среди дышащих жабрами гастропод, которые напротив, обитают почти исключительно в них, причём эдемизм этот весьма высокого ранга: из 4 подсемейств переднежаберных гастропод Thiaridae два эндемичны для Танганьики и оба они не встречаются в ассоциированных водоёмах. В Байкале ситуация несколько отлична. И хотя и тут 2/3 эндемов не принадлежат к лёгочным моллюскам, из эндемичных лёгочных моллюсков 40% приндлежит к Acroloxidae (чашечкам) — почти исключительно палеарктическому семейству, представители которого живут или в крупных озёрах, или в реках на быстром течении. Зато эндемичных прудовиков в Байкале нет. В целом 2/3 фауны лёгочных моллюском приурочено к Голарктике (Рис. 4.39).
Возможно, не последнюю роль тут сыграло существенное обеднение биот континентальных водоёмов Голарктики в результате оледенений. На это указывает сравнение числа видов наяд и речных раков североамериканской и евразийской частей Голарктики: в евразийской части обитает 92 вида наяд и 38 видов речных раков, тогда как в североамериканской — 302 и 382 соответственно (Crandall, Buhay, 2007; Bogan, 2007), причём это разнообразие обеспечивается почти исключительно бассейном Миссисипи, не подвергавшемся оледенению. Сходную картину дают и другие группы, например пресноводные рыбы (Рис. 4.43), лишь крабы разнообразнее в Палеарктике, нежели в Голарктике, что, вероятно, связано с их крайне высоким разнообразием в тропической Азии (Рис. 4.36).
На снижение роли ценофилов в умеренных районах Северного полушария указывает и то, что, хотя максимальные размеры рыб здесь те же, что и в Африке, Южной Америке и Юго-Восточной Азии, мелкие рыбы, весьма разнообразные в низких широтах, в Голарктике практически отсутствуют. Всё это снижает биологический смысл районирования, проведенного методом сравнения списков биот.
Высока в умеренных районах роль и периферических таксонов, в частности проходных и полупроходных рыб (лососи Salmonidae в Северном полушарии и родственные им галаксиды Galaxiidae в Южном). Причиной этого, вероятно, является отсутствие или снижение разнообразия многих групп водных животных, в первую очередь облигатно пресноводных.
Голарктике свойствен также такой не имеющий аналогов в других областях таксон как бобры, эффективно перестраивающий систему водоёмов.
Биоты южных районов областей Южного полушария включают некоторое количество первично пресноводных таксонов. Таковы изоподы подотряда Pheatoicidea, бокоплавы Neoniphragidae и Paramelitidae, речные раки Parastacidae. Это — облигатно пресноводные таксоны, способные расселяться исключительно по пресным водоёмам, поэтому они несомненно возникли в то время, когда существовала такая возможность, т. е. по крайней мере до разделения Гондваны. Иногда их ареал охватывает тропические районы, но обычно в тропиках расположены рефугиумы (Phreatoicidea в Индостане). Лишь при отсутствии конкуренции они проникают в тропики, как это обстоит в Австралии, лишённой многих облигатно пресноводных таксонов. Для ареалов такого типа была предложена гипотеза оттеснённых реликтов. В данном случае эту гипотезу следует слегка модифицировать, предложив гипотезу их вымирания в тропических районах. Вероятно они вымерли в ареале тропических биот потому, что не смогли войти в состав сформировавшихся позднее тропических ЦС. Северная граница их ареалов совпадает с наиболее значимыми биогеографическими границами данной области. Гипотеза оттеснённых реликтов хорошо объясняет, почему ареалы таких таксонов охватывают или острова (Мадагаскар, Новая Зеландия, Фолкленды) или умеренные биогеографические регионы на материках (провинция в Эфиопской и подобласти в Австралийской и Южноамериканской областях). P. Banarescu предложил другую гипотезу, согласно которое такое распространение обусловлено изначальным предпочтением ими умеренных районов, что выглядит гораздо менее правдоподобным и меньше согласуется с фактами.
В то же время ограниченность ареалов многих пресноводных таксонов не имеет сколь-нибудь приемлемого объяснения, например ареалы крабоидов Aegidae (Рис. 4.23) и жемчужниц (Рис. 4.29).
Существенное отличие биогеографии пресных вод от наземной биогеографии — наличие неизолированных регионов высокого эндемизма. Прежде всего это относится к озёрам, особенно крупным. Их биоты резко отличны от биот окружающих водоёмов, что заставляет рассматривать такие озёра как самостоятельные биогеографически выделы, порой очень высокого ранга. Внутри озера формируется своя ЦС, а, если озеро достаточно велико, то и несколько ЦС. Поскольку озёрные биоты являются производными местных биот, даже высокоэндемичные биоты наследуют и их особенности. В частности, для озёр районов с умеренным климатом характерно разнообразие бокоплавов (Байкал, Титикака) и изопод (Байкал), но не рыб, тогда как для тропических (например, в Великих Африканских озёрах) — наоборот, разнообразие бокоплавов и изопод мало, а рыб — высоко. Даже при сравнении озёр гораздо меньшего размера и с гораздо меньшим эндемизмом, расположенных в разных климатических поясах (Бива — в субтропическом и Ланао — в тропическом) мы наблюдаем ту же закономерность.
Основные отличия биогеографии континентальных водоёмов и суши суммированы в Табл. 4.10.
Табл. 4.10. Некоторые различия биогеографии континентальных водоёмов и суши.
|
суша |
континентальные водоёмы |
|
|
растительность |
основа районирования |
малоинформативна |
|
широтная зональность |
проявляется в чередовании типов растительности |
отсутствует |
|
регионы высокого эндемизма внутри других |
отсутствуют |
имеются (озёра) |
|
доля ценофобов в биоте |
<1/2 |
обычно ”1 /2 |
|
видовое богатство к экватору |
увеличивается почти во всех крупных таксонах |
в одних крупных таксонах увеличивается, в других — уменьшается |
|
регионы максимального видового богатства ценофилов |
тропики |
тропики и бассейн Миссисипи |
|
различия Ориентального, Африканского и Мадагаскарского регионов |
сравнительно невелики (ранга подобластей) |
весьма существенны (ранга областей) |
|
Новая Гвинея |
часть Палеотрописа |
часть Австралийской области |
|
древние таксоны с постоянно узкими ареалами |
отсутствуют |
имеются |