Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Голарктика - Раздел 2 - Схемы биогеографического районирования - Биогеография континентальных водоёмов
Районы с умеренным климатом в Южном полушарии в пределах каждого крупного массива суши, вероятно из-за их сравнительно небольших размеров, населены сравнительно однородной биотой. В Северном полушарии умеренные районы занимают площадь, превосходящую на несколько порядков площадь всех умеренных районов Южного полушария вместе взятых. Это позволило сформироваться не просто собственной умеренной биоте Северного полушария, но биоте, существенно различающейся в разных районах, вполне заслуживающих выделения их в отдельные подобласти, а в совокупности дающую биоту, заслуживающую выделения в отдельную область.
Границы области разные авторы проводят по-разному (Рис. 4.9—4.12, 4.30, Рис. Ц7, 9), особенно велики различия в положении границ на Дальнем Востоке. Где проводить границу между Голарктикой и Ориентальной областью остаётся неясным до решения концептуальных вопросов, в частности вопроса о том, на чём надо основывать биогеографическое районирование. Подсчёт преобладания тех или иных географических элементов в регионах, выделенных по географическим признакам имеет все врождённые пороки метода сравнения списков биот и не способны существенно помочь в решении этого вопроса. Бесперспективным являются попытки заменить решение концептуальных вопросов бездумным вычислением разных коэффициентов. Формальные математические процедуры не могут заменить содержания. Точность науки определяет не степень использования математического аппарата, а точность теорий, лежащих в её основе, т. е. непротиворечивость ее исходных посылок, точность и достоверность следствий из них. В этом математика обычно помочь не в состоянии. Как правило, критерием служит “очевидность на качественном уровне” для автора теории и его последователей. Математика может сказать только, КАК считать, но не может сказать, ЧТО считать. Схемы, основанные на механическом вычислении коэффициэнтов обычно способны только вызвать удивление своей неадекватностью. Например, на схеме Четропруда (2010), несмотря на то, что она постулирована как единая для макробентоса, нет какого-либо биогеографического выдела, соответствующего Байкалу.
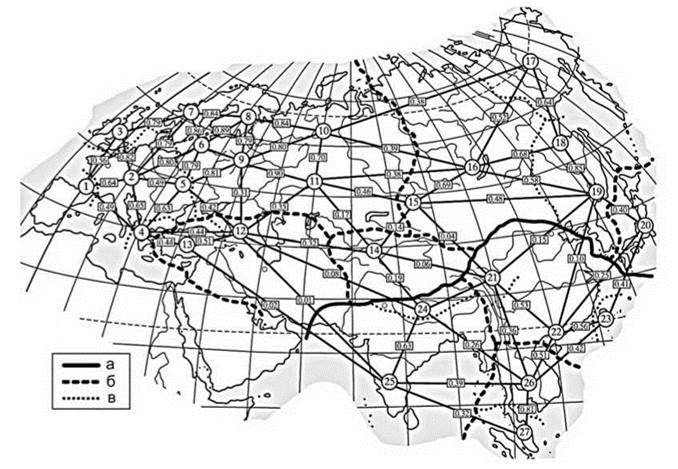
Рис. 4.30. Сходство регионов Евразии по всей фауне макробентоса и обобщенная схема биогеографического районирования. По Чертупруду (2010).
Границы: а — областей, б — подобластей, в — провинций. Цифры в кружках — районы выделенные по произвольно выбранным небиологическим критериям “имеющие в составе горный массив и прилегающие равнинные участки; при прочих равных — более изученные фаунистически и удаленные от соседних регионов” (Чертопруд, 2010: 146) В прямоугольных рамках — значения индекса сходства Хаккера-Дайса. В анализе по неуказанной в статье причине использована только треть фауны рассматриваемых групп.
Из авторов, использующих биологический подход, Л.С. Берг (1939) проводит границу области по южным границам ареалов таксонов, распространённых преимущественно на севере Евразии и Северной Америки. Эти таксоны становятся поэтому её эндемами, что позволяет ему писать, что она “характеризуется распространением исключительно ей свойственных семейств Salmonidae <лососёвые>****, Thymallidae <хариусовые>, Umbridae, Esocidae <щуковые>, Percidae <окунёвые> и др., родов Petromyzon <морские миноги>, Lampetra <речные миноги>, подсем. Scaphirhynchini*****, родов Cottus <бычок-подкаменщик>, Lota <налим> и других” (Берг, 1939: 1238). Понимаемая таким образом область должна была бы включать и бассейн Амура, но он выделяет его в отдельную переходную область. Другие авторы (Старобогатов, 1970; Banarescu, 1992) придают большее значение наличию таксонов, идущих с юга и общему облику биоты. Оба этих автора бассейн Амура (включая ранее также входивший туда Сахалин) относят не к Голарктике, а к Ориентальной области. Более того, P. Banarescu пишет: “водораздел между бассейнами Амура и Лены — одна из наиболее резких границ пресноводных фаун в мире — разделяет Голарктику и Индо-Китайскую область” (Banarescu, 1992: 527). Авторы схем по насекомым: стрекозам (Белышев и Харитонов, 1981) и ручейникам (Moor, Ivanov, 2008), напротив, переходную область Л.С. Берга (1939) относят к Голарктике. Т. е. фактически различия схем состоят в том, к какой из областей относят бассейн Амура и прилежащие территории (Корею, Японию, Китай).
“It is worth mentioning that the water divide between the basin of the Amur and Lena Rivers is one of the clearest boundaries for freshwater faunas in the world, separating the Holarctic from the Sino-Indian region” (Banarescu, 1992: 527)
“Переход щуковых от морского к пресноводному образу жизни, видимо, произошёл в Сарматском море при его опреснении. Щуковидные распространились по пресным водам Лавразии, дав три семейства: щуковых (Escidae), даллиевых (Dalliidae) и Умбровых (Umbridae)” (Микулин, 2003). Мнение о принадлежности австралийских Lepidogalaxias к щуковидным в последнее время оставлено (Leveque et al., 2008).
Независимо от положения границы области на Дальнем Востоке, некоторые таксоны явно приурочены к Голарктике. Из рыб это лососёвые Salmonidae и близкие сиги Coregonidae и хариусы Scaphirhynchinae, отряд щуковидные Esociformes (Esocidae, Umbridae, из которых некоторые выделяют Dalliidae), окунёвые Percidae, бычки Cottidae, некоторые представители очень примитивных семейств рыб, ранее широко распространённые в умеренных водах Голарктики: осетровые Acipinseroidei, ныне исключительно североамериканский надотряд Holostei с двумя семействами: панцирные щуки Lepidosteidae (два рода с 10 видами)****** и ильные рыбы Amiidae (1 вид) и другие. Из беспозвоночных это северные речные раки Astacoidea (Cambaridae и Astacidae), гастроподы Valvatidae, водяные ослики Asellidae и многие другие. Некоторые первично пресноводные таксоны возникли в Голарктике. Таковы Margaritiferidae s. str.*******, возникшие, судя по экстраполяциям на основе анализа ДНК, в самом начале мезозоя (минимум 230 млн. лет назад), т. е. задолго до распада Пангеи. Их ареал всегда был ограничен её северными районами, хотя их климат на протяжении этого времени существенно менялся. Ареал Unionidae охватывает все материки, но почти половина видов (мировая фауна — 621 вид) эндемична для Голарктики. Правда, такое высокое их разнообразие обусловлено исключительным их богатством на востоке Северной Америки (почти 300 видов) (Bogan, 2008).
В Голарктике велико таксономическое разнообразие амфибийных насекомых. Если на суше число видов насекомых в каждом отряде в тропических областях существенно превосходит число видов в Голарктике, то соотношение видового богатства водных насекомых показывает сильно отличную картину: число видов в Голарктике превосходит число видов в любой из остальных областей, каждая из которых включает тропические районы (Рис. 4.38—4.40). В то же время таксономическое разнообразие облигатно пресноводных видов таких групп, как рыбы, десятиногие ракообразные, моллюски невелико. В первую очередь это, вероятно, связано со значительным обеденением биоты в периоды оледенений. В пользу такого предположения говорит то, что биота единственной провинции, на которой оледенения сказались мало — Восточноамериканской провинции — значительно богаче биот всех остальных провинций вместе взятых, её таксономическое разнообразие сравнимо с таксономическим разнообразием тропических районов. Другим существенным фактором является наличие бесчисленных эфемерных водоёмов: родников, ручьёв, речек и речушек, часто антропогенного происхождения.
Весьма любопытен ареал двух основных подсемейств Viviparidae. Viviparinae широко распространены по Голарктике (за исключением запада Северной Америки), тогда как Bellamyinae почти исключительно ориентальный таксон, обитающий также в Африке и Австралии, где он, несомненно, сравнительно недавний вселенец. При этом Viviparidae(представители чисто американского подсемейства Lioplacinae = Campelomatinae) в ископаемом виде известны из Южной Америки, где впоследствии вымерли (Старобогатов, 1970).
Эндемичны для Голарктики и хвостатые земноводные.
Мелких рыб, как в тропиках Южной Америки и Азии, в Голарктике практически нет, но максимальные размеры рыб примерно те же: сом Silurus glanis достигал 5 м длины и 300 кг веса, осетровые достигали ещё более крупных размеров, но они не всю жизнь проводят в пресных водах.
До появления человека в поймах Голарктики в массе селились бобры. Влияние их деятельности на пойменные водоёмы можно оценить на примере р. Унжы. Этот левый приток Волги имеет ширину в межень до 100 м и более, в половодье же заливает пойму шириной несколько километров. Водосборный бассейн покрыт лесами, площадь полей незначительна. В лесах обитают многочисленные бобры, охоты на которых практически нет. Судя по моим личным впечатлениям, после стремительного падения численности населения и полного развала сельскохозяйственного производства, все пойменные сенокосы (их также называют пойменные луга) были заброшены и стали быстро зарастать. Одновременно произошло резкое увеличение плотности поселений бобров в пойме реки. Они перегородили плотинами все мелкие ручьи и в результате уровень грунтовых вод в пойме возрос на метр и более. Проходящие по пойме дороги и сенокосы превратились в малопроходимые болота. Таким образом, вместо отдельных водоёмов (русла, старицы, затоны), чередующихся с сенокосными лугами, пойма реки в норме, вероятно, должна представлять почти сплошное низовое болото, с зеркалами воды, соответствующими руслу, старицам и бобровым прудам, над которым возвышаются прирусловые валы (русла и стариц). Задерживание воды в пойменных водоёмах должно снижать как общую водность реки, так и разность уровней воды в межень и половодье. К аналогичным выводам пришли американские исследователи, изучавшие гидрологическую сеть Среднеатлантических штатов США. Они показали, что в результате деятельности трёх-четырёх поколений поселенцев, построивших мельничные плотины, быстрые реки превратились во множество мелких меандрирующих речушек (Walter, Merritts, 2008). Хотя число мельничных плотин было несомненно меньше, чем бобровых, эти антропогенные изменения всё же дают некоторое представление о масштабах влияния деятельности бобров на речную сеть. Возможно, что роль бобров в естественных условиях не столь существенна. В заповеднике “Кологривский лес” численность бобров на мелких речушках эффективно снижают волки. Нелишне напомнить, что, кроме бобров голарктического рода Castor (их вес редко превышает 20-30 кг), в Северной Америке обитали другого рода бобров — Castoroides, вес которых достигал 200 кг, и названные, вероятно, из-за своих крупных размеров C. ohioensis, эти бобры были истреблены ещё индейцами. Впрочем, свидетельств того, что C. ohioensis строили плотины, нет.
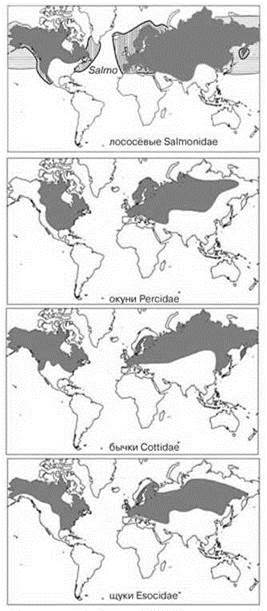
Рис. 4.31. Ареалы некоторых голарктических семейств рыб.
Oncorhynchus — тихоокеанские лососи (горбуша, нерка, кета, чавыча, кижуч и др.), Salmo — настоящие лососи (сёмга, кумжа и др.) Распространение лососёвых в море. По: Савваитова, Медников (1983).
При анализе ареалов голарктических таксонов необходимо помнить о влиянии плейстоценовых оледенений. Во время ледниковых периодов граница Голарктики в Азии и Африки сдвигалась на юг, что привело к возникновению рефугиев на их территории (Абиссинское нагорье, Margaritiferidae в юго-восточной Азии). Одновременно произошло существенное вымирание на обширных площадях. Площади, покрытые ледниками, естественно, полностью лишились своей биоты. Но и не покрывавшиеся ледниками территории в той или иной степени испытали влияние оледенения. В первую очередь оледенение привело к снижению температуры воды и споряжённой миграции биоты на юг. До определённой степени эти миграции облегчало подпруживание рек ледником и развитие обширных приледниковых озёр. В низовьях рек разнообразие ресурсов и биотопов значительно выше, чем в верховьях, поэтому население низовий всегда значительно разнообразнее. Однако на большей части Евразии реки текут с юга на север, поэтому миграции вынужденно совершались из низовьев в верховья. Это привело к катастрофическому вымиранию евразийской пресноводной биоты. Наиболее сильным вымирание было в бассейне Северного Ледовитого океана.
Характерной особенностью районов, подвергшихся влиянию оледенения, является наличие ледниковых реликтов (Рис. 4.32) — эвригалинных таксонов морского происхождения, оставшихся в озёрах после последнего оледенения, когда эти озёра были заливами океана. К этим таксонам относятся бокоплавы Gammaracanthus lacustris, Pontoporeia affinis, Palassea, пять видов мизид рода Mysis (в литературе часто как Mysis relicta) (Audzijonyte, Vainola, 2005, 2007), изопода Mesidotea entomon (крупные озёра Швеции, России и Аляски, этот же вид широко распространён в распреснённых районах умеренных и холодных вод Северного Ледовитого, Атлантического и Тихого океанов), веслоногий рак лимнокалянус, бычок-рогатка Myoxocephalus quadri-cornis labradoricus и нерпы: байкальская и два подвида кольчатой (ладожская (P. hys- pida ladogensis и сайменская Pusa hyspida saimensis, эндемичная для оз. Сайма, два других подвида обитают в Балтийском море и Северном Ледовитом океане). Большая часть этих видов предпочитает низкие температуры и высокое содержание кислорода. Все они имеют близкие виды (подвиды) в Северном Ледовитом океане, Балтийском море, многие и в Каспии.
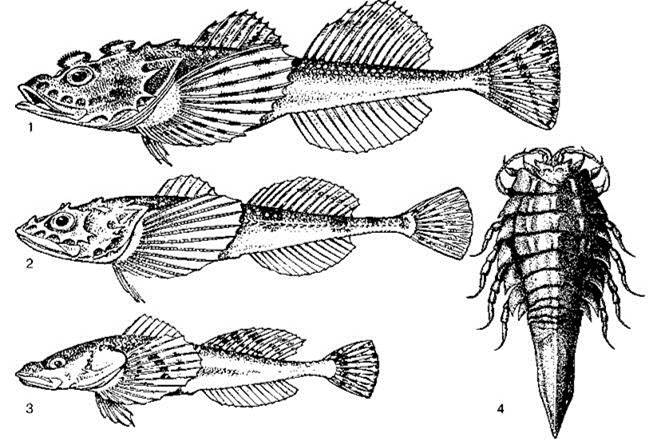
Рис. 4.32. Ледниковые реликты
1—3 — морской четерёхрогий бычок (1) и его озёрные формы: ладожская (2) и онежская (3) рогатки (по Жадин, Герд, 1961), 4 — Mesidotea entomon. По: Kaestner (1959) из Кусакин (1982).
И, наконец, изменение ареалов видов происходило в результате деятельности человека. Это не только многочисленные инвазии (которые при биогеографическом районировании следует игнорировать), но и менее известные случаи. Так, фрагментация ареала Margaritiferidae в Европе произошла уже в историческое время. В частности, на Среднерусской возвышенности существовал промысел речного жемчуга из пресноводной жемчужницы Margaritifera margaritifera. Позднейшее увеличение эрозии в результате сведения лесов и распашки земель привело к резкому увеличению концентрации минеральной взвеси в реках и вымиранию жемчужницы в этой части её ареала. Ныне она сохранилась только в районах, где эрозия из-за особенностей геологического строения местности не может быть столь большой (в частности, в области Скандинавского щита) или где площадь лесов по-прежнему велика.
Голарктика прервана двумя океанами. Атлантический океан широкий, хотя и возраст его не очень велик (палеоцен или ранний эоцен). До этого времени Европа составляла с восточной частью Северной Америки единый континент — Евроамерику. Западная часть Северной Америки была отделена от восточной проливом, существовавшим с раннего мела до раннего палеоцена. С другой стороны с поздней юры до олигоцена Европа была отделена от Сибири эпиконтинентальным Тургайским проливом, располагавшимся примерно в районе Урала. Этот пролив и Паратетис отделяли Европу от большей части Азии (Анатолия была частью Европы). Хотя Тургайский пролив окончательно исчез только в олигоцене, весьма вероятно, что существовал он не непрерывно. В то же время Сибирь с позднего мела составляла единый континент с западной частью Северной Америки (время объединения с южной Азией не вполне ясно). Таким образом, территория нынешней Голарктики, по крайней мере на протяжении части мела, была расположена на двух или трёх материках, очертания которых очень сильно отличались от современных. Такая конфигурация суши привела к значительному сходству раннекайнозойской пресноводной биоты Евроамерики и объясняет распространение некоторых таксонов, например речных раков Astacidae (Рис. 4.24) и веслоносов Polyodontidae (Рис. 4.33) и лопатоносов Scaphirhynchinae (Рис. 4.34).
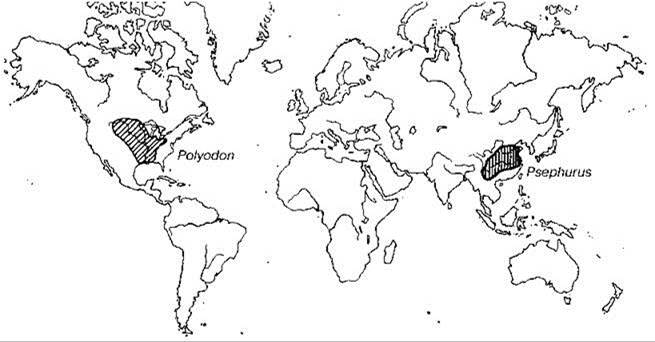
Рис. 4.33 Азиатско-американский тип ареала.
Веслоносы Polyodontidae (Psephurus, вероятно, уже полностью уничтожен). По Banarescu (1990).
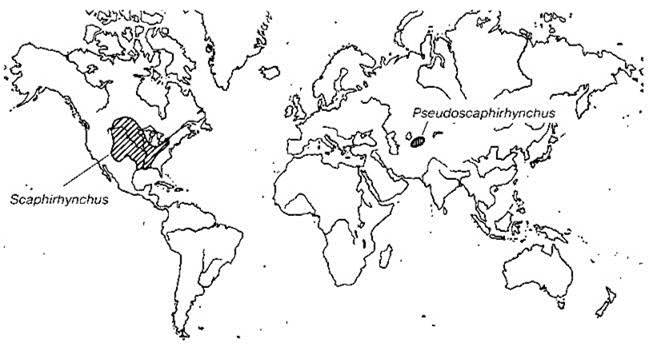
Рис. 4.34 Азиатско-американский тип ареала.
Лопатоносы Scaphirhynchinae По Banarescu (1990).
В то же время, Берингов пролив только в последние 5 млн. лет иногда создавал препятствия для обмена пресноводными биотами Сибири и Северной Америки, по большей же части через сушу, находившуюся на его месте (Берингию) происходил достаточно свободный обмен биотами. Результатом этого обмена являются не только многочисленные ареалы видов, охватывающих крайний северо-восток Сибири и Аляску, но и общность биоты на гораздо более высоком таксономическом уровне. Берингов пролив отсутствовал в периоды, когда температурный градиент экватор-полюс был значительно ниже, а температура в его районе была гораздо выше. Это позволяло мигрировать из Азии в Америку и обратно гораздо более тепловодной фауне, например Cambaridae (Рис. 4.24). В то же время Скалистые горы, возникшие на границе мезозоя и кайнозоя, были гораздо более серьёзной преградой. Поэтому свобода обмена между Азией и Северной, а через неё и Восточной частями Северной Америки была гораздо выше, чем с западом Северной Америки. В пределах Сибири также длительное время существует водораздел Яблоневого и Станового хребта, затрудняющий расселение через него облигатно пресноводных видов. Обмен биот через эти два водораздела путём перехвата верховьев рек был малоинтенсивным и не препятствовал дивергенции биот по их разные стороны.
Голарктика — единственная область, которая имеет границы с тремя областями. Через эти границы происходил обмен. Через границу с Ориентальной областью обмен был интенсивным в обе стороны. Взаимодействие с африканской биотой привело преимущественно к вытеснению её из одних северных районов и обогащением голарктическими видами в других (Нил, Эфиопское нагорье), Unio даже достиг южной оконечности Африки, миграции же африканских видов на север не было. Взаимодействие с Южноамериканской биотой привело к многочисленным обменам периферическими видами (или одновременныму их вселению в оба района), обмена первично пресноводными видами не было совсем.
В предложенных схемах районирования Голарктики существует много различий. Во-первых, одни авторы разделяют Голарктику на Палеарктику и Неарктику (Старобогатов, 1970; Graf, Cummings, 2007), тогда как другие (Banarescu, 1992) этого не делают. Особняком стоит схема районирования по ручейникам (Moor, Ivanov, 2008), в которой территория Голарктики разделена на 7 областей, равных по рангу Африканской, Австралийской и Южноамериканской, при этом ни одна из этих областей не соответствует Палеарктике или Неарктике (Рис. Ц7).
Во-вторых, различно положение южной границы Голарктики (Палеарктики) на востоке Азии. Старобогатов (1970: 198) территорию Китая и бассейн Амура относит к Ориентальной области (две подобласти: Амуро-Японская и Китайская), почему-то указывая при этом, что так же поступал и Л.С. Берг (1939), который в действительности Амуро-Японский регион выделял в отдельную Амурскую переходную область, а не относил к Ориентальной области. Banarescu (1991) проводит границу Голарктики и Ориентальной области так же, как и Я.И. Старобогатов. В то же время авторы схемы районирования по наядам (Graf, Cummings, 2007), напротив, эти территории включают в состав Палеарктики.
Сравнивая приведённые выше схемы (Рис. 4.9-4.12 и Рис. Ц7) можно найти много других отличий. Ниже принята схема районирования Голарктики P. Banarescu (1992) с незначительными изменениями. Он выделяет в Голарктике 8 провинций.
Фаунистически богатейшей является Восточноамериканская провинция (см. также Рис. Ц8). Это связано в первую очередь с тем, что на территории провинции не было оледенений. При похолодании теплолюбивые виды могли отступать по Миссисипи на юг (из верховий в низовья!), а затем возвращаться на север. Поэтому биота провинции пострадала от оледенений гораздо меньше, чем биоты других провинций Голарктики. Большая часть провинции занята водосборным бассейном крупнейшей реки провинции — Миссисипи. Эта река, хотя и не очень полноводна (по среднегодовому расходу воды находится на 8-м месте, уступая многим рекам, в том числе Енисею), но по площади водосборного бассейна занимает третье место. Группа Великих озёр — крупнейшее в мире скопление пресной воды площадью 245 тыс. кв. км и объёмом воды 23 тыс. куб. км (столько же, сколько в Байкале). Климат провинции от умеренного до субтропического. Из рыб наиболее разнообразны карповые, хотя и не столь как в Евразии. Есть и несколько эндемичных семейств, среди которых выделяются весьма примитивные Amiidae с единственным видом Amia calva, ископаемые представители семейства известны из Европы. Зато пресноводные виды лососёвых отсутствуют на большей части территории провинции, в частности в бассейне Миссисипи, встречаясь лишь на севере, проходные виды встречаются шире, но тоже отсутствуют на большей части провинции, в реках бассейна Мексиканского залива и южных реках Атлантического побережья. Очень разнообразны черепахи провинции: 55 видов 16 родов, что вдвое превышает их разнообразие в Африке, включая Мадагаскар (25 видов 6 родов). Весьма разнообразны и беспозвоночные. Unionidae представлены почти 50 родами (почти 300 видами), Ambleminae, представленные тремя эндемичными трибами. Речные раки представлены Cambaridae, образующими 11 эндемичных родов (374 видов) (Crandall, Buhay, 2008).
Биота Западномериканской провинции резко отличается от Восточноамериканской. Общими для них являются только один вид рыбы и несколько видов лёгочных гастропод и нет ни одного общего вида наяд, переднежаберных гастропод и речных раков. Более того, речные раки представлены не Cambaridae, а эндемичным родом Astacidae(Pacifastacus). Unionidae представлены эндемичной монотипичной трибой Gonideini. Фауна провинции гораздо беднее фауны Восточноамериканской провинции, особенно фауна первично пресноводных таксонов. Так, из рыб здесь отсутствуют встречающиеся во всех других провинциях Голарктики окунёвые Percidae, щуки Esocidae, сомы Siluridae(отсутствуют и в Арктической американской). Даже в общих (или викарирующих) таксонах число видов значительно меньше, а ареал таксона часто не охватывает всю провинцию (как у водяных осликов Asellota). Зато пресноводные и проходные виды лососёвых распространены по всей провинции, компенсируя малое разнообразие первично пресноводных рыб.
Фауна Евро-Средиземноморской провинции сравнительно бедна и включает многие широко распространённые по Голарктике или даже Голарктике виды или рода, образующие здесь немногочисленные эндемичные виды. Особо следует отметить осетровых, фауна которых хотя и немногочисленна, но в реках Понто-Каспийского бассейна гораздо более разнообразна, чем где бы то ни было. Речные раки представлены Astacidae.
На территории провинции расположено озеро Охрид (площадь 348 км2, глубина до 285 м), самое древнее озеро Европы, являющееся очагом локального эндемизма и рефугиумом доплейстоценовой биоты. Многие виды рыб, широко распространённые в Голарктике, но отсутствующие на Балканах, сохранились здесь. К тому же специфика жизни в озере, как обычно, способствовала вспышке эндемизма. Для озера характерно наличие эндемов в разных таксонах (некоторые из них выделяют иногда в самостоятельные рода), но особенно — высокий эндемизм гастропод Hydrobioidea, представленных 33 эндемичными видами и 20 эндемичными родами и даже двумя эндемичными подсемействами. Второе по величине озеро Балкан, Prespa (площадь 285 км2, глубина до 54 м), расположенное в 16 км к юго-востоку от оз. Охрид и имеющее в него подземный сток, также является центром эндемизма тех же таксонов, что и Охрид, но гораздо менее выраженным.
Наличие температурного градиента экватор—полюс вызывает возникновение широтной зональности и в пределах провинций. И здесь при прочих равных биоты более южных районов более разнообразны. Увеличение числа таксонов может происходить как путём видообразования, так и благодаря миграциям из прилежащих районов более южных областей. Однако обмен биоты Голарктики с более южными областями в южных районах Северной Америки и Европы был затруднён физико-географическими особенностями этих районов. Аборигенные таксоны по какой-то причине не смогли обеспечить необходимый рост разнообразия биоты. Это привело к тому, что для этих районов Голарктики характерна повышенная доля вторично пресноводных и периферических таксонов. Это характерно для Евро-средиземноморской провинции, но из-за общей её бедности, а также из-за наличия понто-каспийского центра таксонообразования, создающего фаунистический градиент почти в том же направлении, выделение отдельных регионов здесь обычно не проводят (Старобогатов, 1970; Banarescu, 1992). В гораздо большей степени это явление характерно для юга Северной Америки, который P. Banarescu (1992) выделяет в отдельную Центральноамериканскую провинцию, охватывающую как атлантический, так и тихоокеанский склоны материка.
Из-за короткого вегетационного периода продуктивность водоёмов Крайнего Севера и Сибири понижена, в таких условиях не все таксоны могут давать виды, способные поддерживать жизнеспособные популяции. В результате низовья рек бассейна Северного Ледовитого океана беднее, чем их верховья, хотя обычно картина обратная: низовья значительно богаче верховьев. Дополнительная причина обеднения северных районов — голоценовые оледенения, в результате которых вымерла значительная часть биоты, поскольку большинство видов биоты низовьев не могло отступить в верховья, ибо там нет походящих биотопов (ситуация обратная той, что была в Восточноамериканской и даже Средиземноморской провинциях). Та же ситуация была и в реках, текущих в Охотское и Берингово моря, климат которых не мягче климата на побережьях Северного Ледовитого океана. В Беринговом море, кроме того, возможно, образовывался шельфовый ледник. Это привело к существенному вымиранию доплейстоценовой биоты. Ныне в водоёмах Крайнего Севера повышена доля вторично пресноводных таксонов, в частности из рыб доминируют лососёвые.
В Северной Америке таким районом с обеднённой биотой является Арктическая американская провинция. Почти вся территория, занимаемая этой провинцией, во время последнего оледенения была покрыта ледником, поэтому была полностью лишена какой-либо жизни. Граница оледенения примерно совпадает с южной границей провинции, проводимой P. Banarescu (1992). Её биота, за исключением перечисленных выше биогеографических групп, является сильно обеднённой биотой Восточноамериканской провинции. В Евразии таким районом является Сибирская провинция. Её биота также имеет очень незначительное число эндемов и представляет собой сильно обеднённую биоту Евро-средиземноморской провинции, на юго-востоке с примесью видов, идущих из бассейна Амура, а на востоке — из Северной Америки. Из отрицательных черт провинции можно отметить бедность карповыми, Unionidae, Isopoda, а десятиногие ракообразные вовсе отсутствуют. Из карповых в провинции широко распространены только гольян Phoxinus phoxinus, пескарь Gobio gobio, плотва Rutilus rutilus, елец Leuciscus leuciscus, язь L. indus, линь Tinca tinca, карась Carassius carassius, Eupalassela percnura, Orthrias barbatulus. Unionidae представлены только Anodonta (широко — только A. cygnea), Isopoda — только викарирующими видами Asellus. Сибирской провинции принадлежат бассейны рек Азии, впадающие в Северный Ледовитый океан (биота европейских имеет переходный характер), а также в Бергингово и Охотское моря и реки Камчатки.
Сходные с арктическими провинциями условия свойственны Высокогорной провинции, занимающей центр Азии (Гималаи, Тибет, Тянь-Шань и др.). Для этого района характерно подсемейство карповых Schizothoracinae и некоторые рода других семейств, а также полное отсутствие десятиногих ракообразных и Unionidae, бедна и фауна остальных первично пресноводных таксонов. Зато насекомые относительно разнообразны, особенно ручейники. Эту провинцию выделяют в отдельный на всех схемах, но объединяют её то с Голарктикой (Берг, 1939; Старобогатов, 1970), то с Ориентальной областью (Banarescu, 1992). Из-за сложного рельефа района её пресноводные водоёмы часто сильно изолированы. Поэтому многие виды имеют локальные ареалы, ограниченные верховьями тех или иных рек. В то же время, из-за непрерывно идущих горообразовательных процессов, провинции свойственно частое изменение речной сети (перехват верховьев рек), что, наряду со сходством абиотических условий и способствовало сравнительной однородности биоты района. Необходимо также помнить, что вся провинция весьма молода: ещё в эоцене на месте этой крупнейшей на Земле горной системы были моря. Горы возникли лишь 45—25 млн. лет назад в результате присоединения Индостана к Азии. К моменту их возникновения пресноводная биота окружающих районов была весьма разнообразной и многие таксоны оказались способны участвовать в формировании биоты Высокогорной провинции. Это объясняет сравнительно низкий эндемизм её биоты.
На территории Голарктики расположено озеро Байкал. Его биота резко отличается от биот окружающих районов. Как во многих озёрах такого размера здесь образовались свои ЦС, состоящие в значительной степени из эндемов. Кроме того, вымирание сибирской биоты в ледниковые периоды усиливало контраст биоты Байкала и Сибири. Поэтому во всех биогеографических схемах пресных вод Байкал выделяют в отдельный регион ранга равного перечисленным выше провинциям (Берг, 1949; Banarescu, 1992) или всей Голарктике (Старобогатов, 1970). Биогеография озера была рассмотрена выше.
На территории Сибирской провинции имеются и другие районы, для которых свойственен локальный эндемизм. Это или крупные озёра (Зайсан, Иссык-Куль, Балхаш), или верховья рек, или территории, на которых расположены группы озёр и связанные с ними речные системы. В простейшем случае эндемизм озёр связан, как обычно, с локальным видообразованием. Но эндемизм некоторых районов, как например западной Монголии с озёрами Убса-Нор, Киргиз-Нор и верховьями Оби, по-видимому связан с тем, что эти районы также являются и рефугиумами. Берг (1949) выделил западную Монголию в отдельную провинцию, которую он объединил с высокогорноазиатским районом. P. Banarescu (1992) принимает его районы, но объединяет западную Монголию с Сибирью, а высокогорноазиатский регион относит к индокитайской области. Я.И. Старобогатов (1970) выделил в отдельную провинцию чуть ли не каждый бассейн, если в нём имелось хотя бы незначительное число эндемов. По моему мнению, выделение многочисленных регионов для районов, имеющих незначительное число эндемов, на данной стадии развития биогеографии нецелесообразно.
На территории Голарктики расположено известное озеро с повышенным эндемизмом — Бива (Biwa). Площадь озера 670,4 кв. км, глубина 103,58 м. Это озеро находится на Хонсю (Японские острова) около древней столицы Японии — Киото. Озеро воспето в японской культуре, что и обеспечило ему широкую известность. Оно возникло как мелкое маленькое озерко 5—6 млн.л.н., современная глубоководная котловина имеет возраст около 300 тысяч лет. Для него эндемичны 19 видов гастропод (50%) из родов Heterogen(Bellamyinae), Semisulcospira, Radix, Gyraulus, 9 (из 16) двустворчатых моллюсков Anodonta, Corbicula и Pisidium, монотипичные олигохеты Biwadrilinae, эндемичный род пьявок Ancyrobdella, и 6 видов и подвидов рыб (около 10%): сомы Silurus biwaensis и S. lithophilus, бычок Gymnogobius isaza, донно-пелагические Nipponocypris sieboldii, Sarcocheilichthys biwaensis, Squalidus chankaensis biwae, Gna- thopogon caerulescens (Старобогатов, 1970; Banarescu, 1991; Leveque et al., 2008, fishbase; http://www.panda.org/about_our_earth/ecoregions/lake_biwa.cfm). Впрочем, возможно, эндемизм озера сильно преувеличен (см. разбор методов биогеографии).
*В связи с довольно сильным отличием биоты Эфиопии от остальной части Африки название области “Эфиопская” не очень удачно и поэтому здесь заменено на “Африканская”.
** По Я.И. Старобогатову (1970) — отдельное подсемейство с множеством родов.
*** Вероятное число эндемичных видов цихлид оценивают в 800 (Leveque et al., 2008) и даже 1000 (Kornfield, Smith, 2000).
**** Проходные виды лососёвых встречаются и в Индо-Китайской области, в низовьях Нила и в реках Персидского залива.
***** Лопатоносы, современное правильное написание Scaphirhynchinae, окончание -ini ныне используют для триб.
****** Ископаемые остатки современных и близких к ним родов известны из палеоцена Европы, мела Индии и Африки.
******* Я.И. Старобогатов (1970) включал в состав Margaritiferidae много родов из Азии, Европы и Северной Америки, которые он отнёс к подсемействам Arcidopsinae и Pseudodontinae. Эта классификация никем не была признана, а рода отнесены к Unionidae.