Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Моделирование континуальности и дискретности - Континуализм и структурализм - Био-экология, или синэкология
С III Ботанического конгресса в Брюсселе (1910 г.) экологию принято разделять на аутоэкологию и синэкологию. Эти две части биологии, хотя и образованы от одного слова “экология”, имеют мало общего, отличаясь как предметом, так и методами. Синэкология изучает экосистемы, их структуру и функционирование. Аутоэкология изучает биологию отдельных видов, т. е. имеет тот же объект, что и таксономия и морфология, поэтому её логичнее считать частью систематики*.
Хотя каждый вид животных и растений обитает совместно со многими видами, многие современные экологи ограничиваются изучением аутэкологии, т. е. одного, двух или немногих видов, фактически оставаясь почти целиком в рамках систематики, а не экологии (синэкологии). В результате появились такие широко используемые словосочетания “экология растений”, “экология животных”, “экология насекомых”, крайним вариантом такого словоупотребления являются выражения типа “экология вида” или “популяционная экология”, “демэкология” и даже “экология особи”. Впрочем, встречаются гораздо более противоестественные словосочетания, например “молекулярная экология” (Freeland, 2006). В учебниках по экологии основное место уделяют динамике популяций и их взаимодействию (при том, что популяция не менее неопределённое понятие, чем экосистема). Фактически они превратились в интересные, но бессистемные рассказы об экосистемах. Отдельные главы посвящают конкуренции, экологической нише и т. п. И в этом направлении сделано очень много.
“...экологией мы будем называть то, чем занимаются экологи” (Гиляров, 1992: 6).
“Все стали “экологами”. Такого взрыва профанации знания не было в истории человечества” (Реймерс, 1992: 13).
“Под экологической системой мы будем понимать не только “экосистему” в собственном смысле этого слова, но и популяции, гильдии, сообщества, т. е. любые надорганизменные объекты, изучаемые экологией” (Гиляров, 1988: 204).
“Не имеет значения, как мы вычленяем исследуемую часть экосистемы. Часто удобными оказываются естественные границы... или административные, например границы города,.. они могут быть и условными” (Одум, 1986: 28).
“The size or scale varies according to the interests and needs of the researcher. An ecosystem can be as small as a 10 cm... or the Earth itself” (Woodward, 2009: 53)
“Сообщество — “некая единица живой природы, которую можно охарактеризовать в соответствии с признаками, представляющими для нас интерес” (Бигон и др., 1989: 113).
Однако в изучении самих экосистем прогресс не столь значителен. Если в физике нет разных мнений о том, что такое атом, а в химии — молекула, то в экологии ситуация существенно отлична. Экосистемами (биоценозами, биогеоценозами, сообществами) называют любые структуры надвидового уровня, на которые разделяют биосферу. Их смысл столь сильно перекрывается, что, определив любой, мы определим все остальные. И, хотя определений существует множество, все они довольно расплывчаты и очень субъективны. Это привело к тому, что в настоящее время экосистемы выделяют не на основе свойств изучаемого объекта, а по каким-либо произвольным признакам, например, по ландшафтам, почвам, грунтам и т. п., иногда даже вообще без каких-либо критериев, т. е. субъективно. На практике экосистемой называют население произвольно выделенной в соответствии с задачами исследования части поверхности Земли, которую удаётся обследовать имеющимися ресурсами (людскими и материальными). Фактически единственное “конструктивное”, т. е. применимое на практике определение гласит: экосистема (биоценоз и т. п.) — это то, что изучает в текущий момент эколог, если он не утверждает обратного. Или, говоря более наукообразно: “Экосистема есть отображение в сознании исследователя тех свойств объективной реальности, которые нужны ему для решения поставленной задачи” (Шитиков и др., 2003).
“Каждая устричная банка в определённой степени представляет собой сообщество живых существ, подборку видов и определённое количество индивидуумов, которые как раз на этом месте находят все условия для своего проявления и жизни, т. е. соответствующую почву, достаточное питание, надлежащее содержание солей и подходящую, благоприятную для своего развития температуру... Я предлагаю название “Biocenosis” для такого сообщества (Mobius, 1877, цит. по Кафанов, 2005: 27—28).
“The list of evolutionary and ecological “rules” and “laws” is brief, and primarily of historical rather than current interest” (Gaston et al., 1998: 70).
“Our understanding of the organizational structure and dynamical behavior of the systems that contain... organisms is still rudimentary” (Brown, 1999: 4).
“Всем известно, что единорог — существо иного мира и предвещает счастье, — об этом говорят оды, труды историков, биографии знаменитых мужей и другие источники, чей авторитет бесспорен. Даже дети и простолюдины знают, что единорог сулит удачу. Но зверь этот... с трудом поддаётся описанию... И потому, окупавшись перед единорогом, мы можем его не узнать” (Борхес, 1994б: 89).
“...students of ecology will continue to be trained primarily as botanists, zoologists, sociologists, or economists for some time to come — probably indeed as long as university departments are organized on the present basis. Hence, to emphasize the proper synthetic approach and to maintain the ideal constantly before specialized workers, the term bio-ecology appears to be well warranted” (Clements, Shelford, 1939: 1—2).
“Части <системы> заменяемы... отграничение системы обусловлено внешними условиями, а не внутренними предпосылками” (Тишлер, 1971: 11).
Итак, для простоты экосистемой называют всё что угодно, но проще ситуация от этого не становится. Конечно, выработку единой точки зрения по ряду принципиальных вопросов организации природных экосистем затрудняет необычайное их разнообразие, но это не объясняет, почему прогресс в познании фундаментальных закономерностей экологи столь мал.
Экосистемой называют и гнилой пень, и биосферу. Очевидно, что объекты столь разного масштаба наряду с общими свойствами имеют очень много различий в своей структуре и функциях. Однако проблему выделения экосистемы, как элементарной ячейки биосферы, обычно даже не ставят или вовсе отрицают реальность экосистем (продолжая при этом заниматься экологией!), приводимые в монографиях, руководствах и учебниках определения экосистемы невозможно применить на практике. Отсутствие объективных критериев для выделения изучаемого объекта делает неясным, закономерности чего выявлены в результате исследований, в какой степени эти закономерности являются общими, а в какой — специфичны для изучаемых объектов. Сообщества (экосистемы, биоценозы и т. п.) рассматривают в произвольном, но всегда малом масштабе. Независимо рассматривают динамику сообществ, причём, поскольку сообщества выделяют произвольно, остаётся неясным, динамику чего рассматривают авторы работ. В результате понимание биоценоза (экосистемы, сообщества и т. п.) недалеко ушло от взглядов Мёбиуса полуторовековой давности, согласно которым биоценоз — это “подборка видов”, соответствующая условиям биотопа. Большая часть учебников по экологии отражает существующее положение и заканчивается там, где они должны начинаться: разграничения экосистем в пространстве.
В то же время очень мало кто пытается выявить и изучать эволюционно сформировавшиеся коадаптированные группы видов, многие даже отрицают саму возможность их существования.
Континуализм и структурализм
Среди множества принципиальных вопросов синэкологии один является, безусловно, основным: есть ли вообще в природе экосистемы, или это понятие, придуманное для удобства? Множество ответов на данный вопрос можно расположить вдоль градиента, на концах которого расположены две альтернативные концепции экосистемы. Распределение точек зрения по этому градиенту резко бимодально, что позволяет говорить о существовании в экологии двух парадигм: континуализма (редукционизма) и структурализма (см., например, Миркин и др., 2002). В англоязычной литературе они получили, соответственно, названия: индивидуалистическая гипотеза (individualisticconcept) и гипотеза сообществ-единиц (community units).
“Континуум — фундаментальная черта организации биосферы” (Воронов и др., 2002: 96).
“Континуум — фундаментальная черта организации биосферы” (Абдурахманов и др., 2003: 121).
“Виды встречаются вместе не потому, что приспособились друг к другу, а потому, что приспособились к общей среде” (Миркин, 1985: 51).
“Для континуалиста (Раменский, 1910, 1938; Gleason, 1917, 1939) ценоз — это условно однородный отрезок континуума, взаимоотношения компонентов в котором не имеют значения; каждый вид — экологическая индивидуальность, по-своему реагирующая на изменения каждого экологического фактора и имеющая по отношению к нему свои показатели обилия” (Кафанов, 2005: 45).
“Трудно отрицать наличие полной взаимонезависимости отдельных видов растений даже внутри сообщества... последней классификационной единицей растительных сообществ являются не ассоциации, а виды растений” (Вальтер, 1968: 12-13).
Парадигма континуализма была впервые сформулирована советским геоботаником Л.Г. Раменским (1924) и независимо от него американским геоботаником H.A. Gleason(1926). В настоящий момент она является доминирующей как в нашей стране, так и за рубежом. С позиций континуализма написаны практически все учебники экологии и биогеографии. Согласно этой парадигме, как её понимают наиболее последовательные сторонники, виды эволюционируют более или менее независимо, воздействия видов друг на друга в сообществе, как правило, неспецифичны. Виды образуют сочетания случайным образом, в зависимости от локальных абиотических условий. Такие случайные сочетания видов — экосистемы, биоценозы, биогеоценозы, сообщества и т. п. — в природе плавно переходят одно в другое, образуя континуум, заполняющий неоднородную среду, как вода в луже заполняет неровности на поверхности земли. Некоторые исследователи даже считают континуальность основным свойством биосферы. Если всё же обнаруживают более или менее резкие границы сообществ, то их всегда считают созданными не биотой, а внешними факторами, например, резкими физико-географическим градиентами. Пример: классический объект, рассматриваемый в качестве экосистемы, — озеро. Фактически это означает признание только двух уровней организации жизни: организм (вид, популяция) и биосфера, а также отрицание существования каких-либо самоорганизующихся биологических структур уровня, промежуточного между видом и биосферой.

Рис. 1.1. Леонтий Григорьевич Раменский (1884-1953).

Рис. 1.2. Henry Allan Gleason (1882-1975).
Отрицая реальность экосистем, континуалисты сводят экологию к изучению популяций и их взаимодействию. Это аналогично попыткам понять разум через изучение строения нервных клеток.
Альтернативная парадигма была впервые сформулирована американским геоботаником F.E. Clements. В 1910-х гг. он работал в университете штата Миннесота, в 1917-1941 гг. руководил экологическими исследованиями в Институте Карнеги в Вашингтоне, автор терминов “сукцессия”, “климакс”, “биом”, “экотон”. В 1970-80-х гг. парадигма была существенно переработана и развита советским геоботаником С.М. Разумовским и его последователями. Она не имеет общепринятого названия. Согласно этой парадигме, биосфера обладает эндогенной структурой, поэтому мне кажется наиболее правильным назвать её структурализм**. Дискретные структуры, из которых, согласно структурализму, состоит биосфера, имеют отчётливые границы, проявляют функциональную активность, направленную на поддержание своей целостности, и эволюционируют как единое целое. Виды в процессе эволюции не приспосабливаются к абстрактной “внешней среде”, а становятся элементами формирующейся экосистемы, функционирующей в определённом диапазоне факторов внешней среды. Ответ экосистемы на внешние воздействия зависит от стадии сукцессии, т. е. может быть весьма различным.

Рис. 1.3. Frederic Edward Clements (1874-1945).

Рис. 1.4. Victor Ernest Shelford (1877-1968).
Сторонники структурализма не только обосновывают свою точку зрения, но и объясняют, откуда берётся континуум в работах континуалистов. К сожалению, конструктивного диалога между сторонниками этих двух парадигм не происходит. Критика континуалистов сводится, в лучшем случае, к отрицанию отдельных фактов и положений структуралистов, всегда в самом мелком пространственно-временном масштабе, где различия между ними не столь очевидны и не всегда существенны. Обычно же о существовании противоположной точки зрения даже не вспоминают, а если и упоминают, то нередко с существенными её искажениями, приписыванием ей положений и высказываний, на ошибочность которых структуралисты сами прямо указывали. Так, например, Б.М. Миркин с соавт. (2002), в таблице 13 сравнивают представления о сукцессии континуалистов и структуралистов по 6 пунктам. При этом сравнивают современные представления континуалистов и взгляды основоположника структуралистов F. Clements (начало прошлого века). Если же сравнивать современные представления и тех, и других, то три из шести приписываемых в этой таблице структуралистам представлений прямо объявил неверными ещё С.М. Разумовский (1981). В результате понять суть концепции структурализма по работам континуалистов столь же сложно, как, например, суть концепции Поппера по учебнику марксистско-ленинской философии.

Рис. 1.5. Станислав Михайлович Разумовский (1929-1983).
Нечёткость понятий “экосистема”, “биоценоз”, “биогеоценоз”, “сообщество” вполне приемлемы в рамках парадигмы континуализма, поскольку задачи исследований и не требуют такой чёткости (ибо приверженцы этих взглядов априорно отвергают возможность существования самоорганизующихся единиц биосферы). В рамках же структурализма, признающего наличие таких структур, четкость определений существенна.
Обе концепции существуют с начала века, пользуясь переменной популярностью в зависимости от господствующей моды и научной школы, определяемых не в последнюю очередь объектом исследования и используемыми методами. И континуалисты, и структуралисты основывают свои позиции на фактических данных, полученных в результате исследования природных сообществ.
В тех случаях, когда изучаемая экосистема — достаточно чёткая консорция, или же можно легко выделить виды-эдификаторы, или же изучаемая часть была отграничена уж очень резким градиентом абиоты (например, озеро от окружающего леса), исследователи приходили к выводу о том, что изучаемое ими сообщество — функциональное единство (биотически контролируемое сообщество). Именно лёгкостью проведения границ в значительной степени можно объяснить большую популярность изучения таких нетипичных экосистем, как острова или маленькие озёра. Если же консортные связи, виды-эдификаторы или границы выявить было не столь легко, почти неизбежно следовал вывод о том, что изучаемое сообщество — математический артефакт (физически контролируемое сообщество) (обзор применительно к бентосу см. Несис, 1977).
Континуализм сформирован в значительной степени западноевропейскими и североамериканскими геоботаниками преимущественно второй половины XX в. В связи с этим надо отметить, что на суше, по-видимому, уже тысячелетия отсутствуют ненарушенные, т. е. естественные, не изменённые вмешательством человека сообщества. О том, как они выглядели, можно строить более или менее правдоподобные гипотезы. Диапазон этих гипотез весьма широк.
На одном краю находится гипотеза, согласно которой лесные массивы, не подвергавшиеся сплошной рубке несколько сотен лет (т. е. в несколько раз меньше максимальной продолжительности жизни видов-эдификаторов), являются естественными ненарушенными сообществами.
На другом краю находится гипотеза, согласно которой такие леса необходимо рассматривать как антропогенные промышленные древостои, стоящие гораздо ближе по своим свойствам к сельскохозяйственным полям, и они уж, безусловно, не являются естественными ненарушенными сообществами. Всё большее признание получает точка зрения, что и сами сплошные пояса бореальных (тайга) и неморальных (широколиственные) лесов, а также степи, тундры и многие пустыни имеют антропогенное происхождение. Согласно этой точке зрения, такие пояса возникли на месте экосистем, эдификаторами которых были крупные травоядные (мамонты и другие хоботные, носороги, парно- и непарнокопытные) (Восточно-европейские леса..., 2004) (подробнее см. Главу 3).
То, какой парадигмы придерживается исследователь, в значительной степени определяет его подход к изучению проблемы, методику отбора материала, его анализ, и, следовательно, результаты. Так, широко применяемая при отборе проб практика игнорирования дискретности, часто заметной невооружённым глазом (закладка случайно расположенных площадок, несоотношение размера площадок с мозаичностью, т. е. обычно слишком крупный их размер и т. д.), приводит к тому, что существенная часть площадок захватывает пространства, занятые несколькими сообществами, принадлежащих к разным ассоциациям. Математическая обработка такого неадекватно собранного материала с неизбежностью даёт картину континуума, независимо от действительной ситуации.
“Сама методика случайной выборки, основанная на убеждении об отсутствии в природе объективных границ, никогда не сможет их выявить. Эта методика по своей сути заранее предопределяет результаты исследования” (Киселева, Разумовский, Расницын, 1969: 130).
Представление о том, что для характеристики биоты оптимальным является случайный отбор проб основано на априорном отрицании внутренней структуры биосферы, поскольку с математической позиции такой отбор проб корректен только из одной генеральной совокупности, т. е. из континуума. Если же имеются дискретные структуры, то каждая из них является генеральной совокупностью, что делает неприемлемым случайный отбор проб.
Существенное значение имеет и выбор анализируемых показателей. Если анализировать распределение видов по существенным для структуры факторам, то вполне возможно получить чёткую дискретность. Однако на практике выявить такие факторы обычно очень сложно и анализ проводят по факторам, которые кажутся исследователю значимыми или по которым удалось собрать информацию. Отсутствие дискретности при таком анализе рассматривают как доказательство отсутствия дискретности в принципе, хотя фактически это означает только то, что исследование распределения видов по анализируемым факторам не выявило дискретности.
В изучении водных экосистем случайный (часто нерепрезентативный) характер расположения проб до некоторой степени компенсирует слабая мозаичность донных сообществ рыхлых грунтов и пелагиали.
Поскольку выбранная методология априорно соответствует предполагаемой природе исследуемого объекта, неудивительно, что фактические материалы, собранные и обработанные континуалистами, “объективно доказывают” существование континуума. Поразительным было бы иное. Неудивительно и то, что изучение тех же самых экосистем структуралистами даёт прямо противоположный результат.
В качестве примера рассмотрим интерпретацию данных по распределению видов континуалистами (Рис. 1.6) и структуралистами (Рис. 1.7). Первый рисунок (Рис. 1.6) с незначительными вариациями (иногда дают только один рисунок или наоборот, несколько) широко воспроизводят при обсуждении различий двух парадигм как экспериментальное подтверждение наличия континуума и отсутствия дискретных сообществ (см. например, рисунки в: Одум, 1975: 189; Уиттекер, 1980: 123 и 125; Бигон и др., 1989: 124-125; Brown, Lomolino, 1998: 105; Петров, 2001: 113; Воронов и др., 2002: 97; Cox, Moore, 2003: 52, 73; Нинбург, 2005: 6; Бурковский, 2006: 158 и др.). Сравнение рисунков демонстрирует различия в подходах, осмыслении интерпретации результатов. Главное отличие данных рисунков — не распределение кривых по оси абсцисс, как видится континуалистам, а то, что отложено по оси абсцисс. Континуалисты анализируют распределение видов по градиенту фактора, который они считают значимым, без какой-либо привязки к местности. Такой подход логичен при изучении биотопов уже выделенных сообществ. Однако реальные сообщества (экосистемы и т. п.) занимают место не в воображаемом гиперпространстве факторов, а в обыкновенном вполне реальном географическом пространстве. Пространство факторов — виртуальное, пространством оно является с математической точки зрения, с географической же это просто набор факторов, в котором можно выделить виртуальные группировки организмов. Эти группировки тоже называют сообществами. Свойства реального и виртуального пространств различны. Основная разница состоит в том, что в виртуальном пространстве факторов возможно нахождение в одной точке любого числа видов и сообществ, в географическом же — только одного. Таким образом, происходит подмена понятий, и рисунок, призванный иллюстрировать различия взглядов представителей неодинаковых парадигм на распределение видов, в действительности иллюстрирует разницу в подходах к сбору и анализу данных. В том случае, когда ось абсцисс — градиент фактора результаты вовсе не относятся к обсуждаемому вопросу — к фактическому распределению видов по поверхности Земли и, естественно, никак не могут служить подтверждением справедливости концепции континуализма. Графики, подобные изображённому на Рис. 1.6, показывают распределение видов и их групп по градиенту изучаемых факторов, т. е. их биологические (аутоэкологические) характеристики, но никоим образом не проливают свет на вопрос о степени дискретности реальных сообществ на местности. Методы, используемые континуалистами для выделения сообществ (классификация и ординация), с точки зрения структуралистов в действительности вовсе не являются методами выделения экосистем, а лишь методами выявления групп видов, сходно реагирующих на факторы внешней среды.
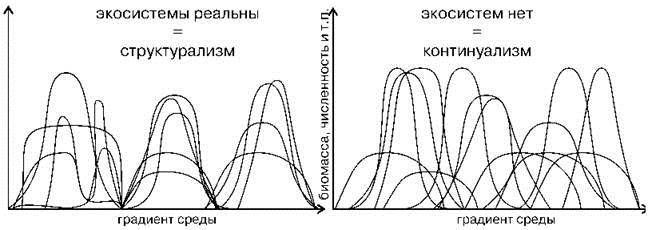
Рис. 1.6. Различия структурализма и континуализма в представлении континуалистов (каждому виду соответствует своя кривая).
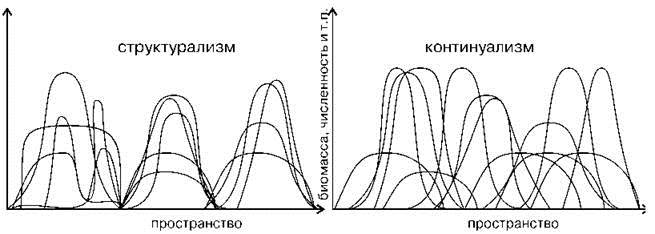
Рис. 1.7. Различия структурализма и континуализма в представлении структуралистов (каждому виду соответствует своя кривая).
Данная идея многим покажется крамольной и абсолютно неприемлемой, поскольку принято считать наоборот. Поэтому давайте применим этот метод (случайных проб и анализа их в пространстве факторов) для изучения территории, несомненно, разделённой в пространстве резкими границами на участки. Например, территорию Европы, поделённую на страны. На территории каждой из стран преобладают её подданные, поэтому будем считать территорию, на которой преобладают подданные одной страны, частью данной страны. Попробуем провести границы стран методами континуализма. Для этого подсчитаем число подданных разных стран в случайно расположенных квадратах, размером, например 50x50 миль, использованных для биогеографического районирования Северной Америки (Hagmeier, Stults, 1964). Очевидно, что часть таких квадратов попадёт на границы и захватит несколько стран. Уже только это снизит резкость границ. Рассмотрим далее распределение подданных разных стран в пространстве любых факторов, например, таких, несомненно, важных для экономики и населения этих стран, факторов, как увлажнение и температура. Даже и без проведения расчётов совершенно очевидно, что мы не сможем получить чётких границ между странами, вместо этого мы получим континуум. А между тем границы такие существуют и необычайно резки (не имеют ширины в изучаемом масштабе). Задача разграничения государств вполне аналогична задаче разграничения сообществ, поскольку и те, и другие занимают какую-то часть поверхности Земли. Становится ясным, что выбранные факторы не способны выявить дискретность. В зависимости от выбранных показателей, мы будем получать более или менее плавный континуум, но никогда не получим чётких границ. Проблема заключается в том, что исследователь, изначально считающий, что дискретности нет, после первого анализа (по увлажнению и температуре) объявит, что математический анализ данных свидетельствует об отсутствии дискретности и наличии континуума, между тем как математически доказано только отсутствие дискретности в распределении подданных той или иной страны в пространстве анализируемых факторов. Сколь бы чёткими ни были границы стран, анализ факторов, несущественных для возникновения этих границ, неизбежно даст нам в результате не чёткие границы, а континуум. И проблема здесь не в выборе адекватного показателя, а в принципиальной неверности самого подхода. Выявить существенные факторы (которыми могут быть и биотические взаимоотношения) удаётся далеко не всегда. Невыявление дискретности в гиперпространстве факторов не может служить доказательством отсутствия чётких границ. Появление континуума в результате такого анализа вовсе не отменит реальности границ, что легко проверить на пункте паспортно-визового контроля, попытавшись вместо предъявления визы, рассказать пограничникам теорию о континууме в гиперпространстве факторов.
“Наблюдения подтверждают принцип Раменского и Глизона о существовании континуума сообществ: широкие перекрытия экологических амплитуд и рассредоточенность центров распределения популяций вдоль градиента среды приводят к тому, что большинство сообществ непрерывно переходят одно в другое, и это случается гораздо чаще, чем образование определённых ясно разграниченных друг от друга сообществ” (Уиттекер, 1980: 125).
“Факты искажают отображение действительности в нашем сознании, если идея, их группирующая, порочна” (Фёдоров, 1987, цит. по 2004: 340).
Таким образом, мнение континуалистов о том, что виды в нормальных условиях образуют не взаимно адаптированные комплексы, а континуум, в котором разные виды в зависимости от факторов внешней среды образуют разнообразные случайные сочетания (см., например, Уиттекер, 1980; Миркин, 1984; Миркин, Наумова, 1997 и др.), основанное на изучении распределения видов по градиентам факторов среды, не только не выдерживает никакой критики, но крайне наивно и неожиданно. Тем не менее, представление это парадоксальным образом широко распространено.
Только непосредственное изучение распределения видов по поверхности Земли (в географическом пространстве, а не в пространстве факторов) способно выявить реально существующие сообщества, хотя и здесь есть свои сложности. В частности, необходим учёт сукцессионной динамики. Однако континуалисты и структуралисты вкладывают разный смысл в понятие “сукцессия” (см. ниже), поэтому и учитывать сукцессионную динамику они будут по-разному.
Появление понятия континуума и особенно его широкая популярность в геоботанике и, отчасти, в гидробиологии (Несис, 1977; Миркин, Розенберг, 1979; Миркин, 1987; Погребов, 1982; 1988) в значительной мере обусловлено не только недостаточным учётом сукцессионной динамики (а зачастую — и полным её игнорированием), но и широким распространением в современной экологии математических методов, не учитывающих пространственное распределение видов. Справедливости ради надо сказать, что существуют модификации широко распространённых методов, с помощью которых можно учесть реальное расположение мест отбора проб в пространстве, но они не входят в наиболее популярные пакеты статистики. Исследователи, следуя традициям (моде?), не ставят перед собой цель учесть пространственное распределение изучаемых явлений. Более того, большая часть биологов о существовании методов, учитывающих пространство, и не подозревает и о необходимости учёта пространства не задумывается. Иными словами, устойчивость представлений континуалистов обусловлена в значительной степени априорным игнорированием пространственной компоненты биологических (в данном случае — экологических) взаимодействий.
В последние десятилетия не в силах отрицать очевидное, т. е. существенное влияние на распространение видов биологических факторов: конкуренции, эдификаторной роли и т. п., концепция континуализма претерпела некоторые изменения. О таких изменениях Б.М. Миркин писал: “Суть новых взглядов на континуум заключается в том, что независимость распределений видов более не абсолютизируется, произошла “фитоценологизация континуума” (Миркин, Наумова, 1998). Виды в растительных сообществах занимают не фундаментальные, а реализованные экологические ниши, объём которых регулируется конкуренцией. Влияют на распределения видов и неконкурентные средообразующие взаимоотношения, например, влияние эдификаторов (таких, как ель в бореальных лесах или сфагнум на верховых болотах). По этой причине распределения видов по градиентам среды могут быть не только симметричными колоколовидными, но и асимметричными и даже би- и полимодальными” (Миркин, 2005: 523).
“На одной из своих лекций Давид Гильберт*** сказал:
— Каждый человек имеет некоторый определённый горизонт. Когда он сужается и становится бесконечно малым, то превращается в точку. Тогда человек говорит: “Это моя точка зрения” (Физики шутят, 1993: 44).
Как становится ясным из приведённой цитаты, нового в действительности очень мало: по- прежнему происходит подмена понятий. Виды влияют на распределение друг друга на местности, в реальном пространстве, а не по “градиентам среды”, не в воображаемом пространстве экологических факторов. Что здесь играет большую роль: непонимание сущности спора или нежелание изменить свою точку зрения?
Введение пространства в классические законы экологии меняет их нетривиальным образом.
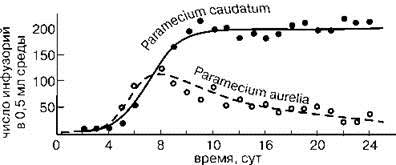
Рис. 1.8. Динамика популяций инфузорий Paramecium aurelia и P. caudatum в совместной культуре По Гаузе (1999) из: Gause (1934).
Так, согласно одному из них — принципу конкурентного исключения — “два вида, занимающие одну и ту же экологическую нишу, не могут устойчиво сосуществовать, поскольку в результате конкуренции один из видов будет вытеснен другим” (Гиляров, 1990: 159). Итальянский математик Вито Вольтерра показал это на системе дифференциальных уравнений. Выпускник кафедры зоологии беспозвоночных биологического факультета МГУ Г.Ф. Гаузе на опытах с инфузориями “в пробирках с плоским дном (диаметр около 1 см и высота 5—6 см) в 0,5 см3 питательного раствора” (Гаузе, 1999: 116), получил экспериментальное подтверждение расчётов Вито Вольтерра. В опыте, результаты которого приведены на Рис. 1.8, при совместном культивировании двух видов Paramecium численность одного росла, а другого падала и через 24 суток после начала опыта (что примерно соответствует 48 генерациям) почти (но всё же не совсем! — ИЖ) упала до нуля. Позднее аналогичные результаты были получены в многочисленных опытах, и этот закон стали считать одним из основных законов экологии, названным по имени первооткрывателей законом (принципом) Вольтерра-Гаузе (Реймерс, 1992; Розенберг, Рянский, 2005). Изучение природы показывает, что сосуществование видов с идентичными потребностями (нишами) — явление достаточно обычное. Например, фитопланктон по всему Мировому океану и в пресных водах состоит из большого числа видов одноклеточных водорослей с очень сходными нишами, причём виды эти встречаются вместе и, по-видимому, сосуществуют многие тысячелетия (если не миллионы лет), но до сих пор друг друга не вытеснили. Данный факт никак не соответствует теории Вольтерра-Гаузе, но тем не менее до сих пор не получил общепринятого объяснения, а лишь название: “планктонный парадокс” (Hutchinson, 1961). Аналогично число видов деревьев, совместно произрастающих в тропическом лесу, очень велико, но в то же время различия их ниш показать не удаётся. Подобные расхождения между теорией и фактами пытаются объяснить двумя способами.
1. Предположением о неполноте фактов:
— поскольку согласно дифференциальным уравнениям Вито Вольтерра достаточно различия в одном измерении ниши, а число измеряемых исследователем в природе параметров всегда конечно, предполагают, что на самом деле виды имеют неодинаковые ниши, просто исследователи эти различия не обнаруживают или же виды ограничены разными ресурсами.
2. Введением дополнительных условий, при соблюдении которых виды с идентичными потребностями всё-таки могут сосуществовать, например:
— неоднородности среды: в опытах Г.Ф. Гаузе, в которых в среду вводили неоднородности (например укрытия), виды с близкой биологией сосуществовали на протяжении всего эксперимента;
— наличие плотностно-зависимых эффектов: ограничение численности разных видов не межвидовыми (конкуренцией за ресурсы, например), а внутривидовыми взаимодействиями. Анализ уравнений (Chesson, 2000; Adler et al., 2007) показывает, что имеются значения коэффициентов, при которых возможно сосуществование видов с идентичными требованиями (нишами).
— конкурентное преимущество меняется по мере расходования (или поступления) лимитирующего ресурса;
— одни виды ингибируют рост других за счет специально выделяемых веществ;
— виды демонстрируют конвергенцию и сосуществование за счет “отложенного вытеснения”;
— между видами есть мутуалистические отношения;
или же привлекается “божественная сила” (вытеснение прерывается нарушениями извне).
Легко заметить, что обсуждение ограничено развитием событий только во времени, хотя реальные события развиваются и в пространстве. Экспериментальные сосуды Г.Ф. Гаузе с точки зрения инфузорий (т. е. учитывая их способность к перемещению в пространстве) равноценны одной точке.
Учёт пространства существенно меняет результат. Проиллюстрируем это наглядной моделью, основанной на идеологии клеточных автоматов**** (Грабовский, 2005).
В отличие от объяснения post factum, на модели легче выделить необходимые и достаточные причины, поскольку в модели условия можно контролировать, а эксперименты — повторять. Такая модель должна:
1. Включать минимум допущений (правило “бритвы Оккама”), которые не противоречили бы имеющимся данным (как палеонтологическим, так и неонтологическим).
2. Демонстрировать поведение системы, которое находится в фокусе наших объяснений.
В жизни, конечно, всё сложнее. Вопрос в том, насколько это принципиально. Ответ на него можно получить, сопоставляя прогноз модели с реальными наблюдениями.
В данной модели особи двух или нескольких видов (число видов можно задать от двух до десяти) взаимодействуют на совершенно однородном моделируемом поле. Особи всех видов имеют равную конкурентоспособность и занимают одну нишу (имеют идентичные потребности). С помощью модели можно изучить динамику во времени: (1) суммарной по полю относительной численности видов и (2) распределение отдельных видов в пространстве. Пространство измеряют в размерах особей, время — в генерациях. Измерения этих параметров в относительных, а не абсолютных величинах (метры, сутки и т. п.) биологически более правильно (Азовский, 2003). При размножении каждая особь делится на задаваемое число эквивалентных ей потомков. Каждый потомок расселяется на случайное расстояние в пределах некоторого задаваемого радиуса. Если выбранный участок оказывается свободным — занимает его. Если участок занят другим видом, то расселяющаяся особь с вероятностью 0,5 (виды имеют равную конкурентоспособность!) либо вытесняет его, либо вымирает. При половом размножении добавляется ещё один задаваемый параметр: радиус поиска полового партнёра (разноса гамет, пыльцы, спермиев, неоплодотворённых яйцеклеток и т. п.). Модель показывает, что виды с одинаковой конкурентоспособностью, занимающие одну нишу в однородном пространстве, могут сосуществовать весьма долго, если это пространство достаточно велико. На модели получено, что продолжительность сосуществования видов уменьшается при:
— уменьшении отношения размера экспериментального поля к радиусу перемещения особей или радиуса поиска полового партнёра (т. е. по мере приближения поля к точке), предельный вариант (поле равно точке) с максимальной неустойчивостью сосуществования видов и рассматривает теория Гаузе-Вольтерра;
— увеличении плодовитости.
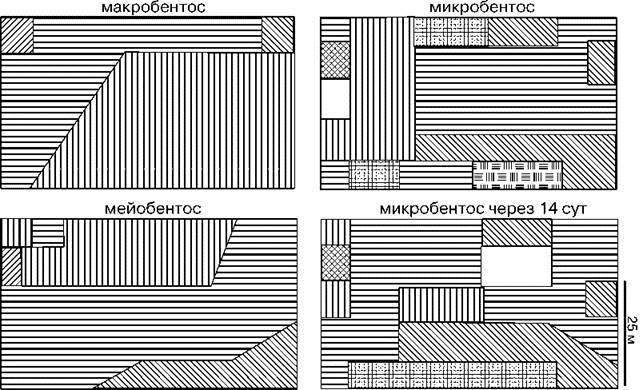
Рис. 1.9. Структурная гетерогенность сообщества на уровне макро-, мейо- и микробентоса.
Тип штриховки соответствует определенной структуре доминирования в микро-, мейо- и макробентосе, выявленной кластерным анализом. Исследованная площадь — 3200 м2, 25 станций (по: Бурковский, 1992). С увеличением размеров организмов возрастает и абсолютный радиус их расселения (в метрах), возрастание абсолютных размеров и радиуса расселения приводит к уменьшению абсолютной мозаичности одновидовых пятен. Динамика положения пятен микробентоса хорошо соответствует прогнозируемым моделью В.И. Грабовского.

Рис. 1.10. Распределение видов в пространстве в том же опыте.
Начальное распределение видов случайное, на рисунке приведено распределение через 100 поколений. Оттенки серого соответствуют тем же видам, что и на рис. 1.10.

Рис. 1.11. Относительная численность десяти видов с идентичными нишами в однородном пространстве через 100 поколений.
Результаты одного из прогонов модели при следующих параметрах: размер экспериментального поля 75 х 75, радиус расселения особей 1, радиус поиска полового партнёра 10, плодовитость 4 потомка, разные оттенки серого соответствуют разным видам). Время эксперимента (100 поколений) примерно в 2,5 раза больше, чем в опыте Г.Ф. Гаузе на Рис. 1.8.
При низкой способности к расселению и большом радиусе поиска полового партнёра (ситуация типичная для, например, деревьев тропических лесов, имеющих крупные плоды, которые разносятся на расстояние гораздо меньшее, чем пыльца), немалое число видов (в данной модели максимальное число — 10) с идентичными нишами способны существовать весьма долго. Прогон модели с параметрами фитопланктона (радиус расселения мал, размножение бесполое) даёт ещё более устойчивую ситуацию сосуществования 10 видов с полностью идентичными экологическими нишами. Виды образуют пятна, форма пятен довольно изменчива, положение пятен более постоянно. Получаемые на моделируемом поле пятна разных видов весьма напоминают наблюдения в природе. Такая картина получается и при изучении распределения сообществ (примерно соответствующему распределению наиболее массовых видов) на литорали Белого моря (Бурковский, 1992, 2006: 154) (Рис. 1.9). На рисунке видно, что, чем крупнее организмы, тем крупнее элементы мозаики. Динамика распределения сообществ микробентоса показывает, что за 2 недели (примерно 30 поколений, т. е. в полтора раза дольше, чем в опытах Г.Ф. Гаузе) форма пятен сильно изменилась, но многие пятна остались примерно в тех же местах. Таким образом, учёт пространства существенно изменяет результаты. При этом получаемые результаты вполне соответствуют наблюдениям в природе не только по моделируемым свойствам, но даже и по тем, какие вовсе никак не учитывали при построении модели. Кроме уже разобранных свойств одновидовых пятен (Рис. 1.10), вполне соответствует наблюдениям в природе и распределение относительных частот видов по результатам модели (Рис. 1.11). Таким образом, оказывается, что противоречие между фактическими данными и обычно используемыми математическими моделями, в частности системой дифференциальных уравнений Вольтерра, объясняется, вероятно, непригодностью выбранного математического аппарата для моделирования таких процессов. Аналогично результаты эксперимента Г.Ф. Гаузе в природе справедливы при соблюдении сделанных при его постановке допущений: особи способны расселяться на расстояние, сопоставимое с размерами однородного участка, в котором происходит взаимодействие видов. Построенная же на основании этих моделей и экспериментов общая теория нуждается в существенной уточнении, поскольку они описывают поведение системы в условиях, почти никогда не наблюдаемых в природе. Поэтому формулировку закона Гаузе-Вольтерра следует уточнить следующим образом: “Если два или более видов, занимающих одну и ту же экологическую нишу, поместить в малый объём, и при этом особи видов способны без затруднения перемещаться по всему объёму, то при ограничении необходимых видам ресурсов, достаточно длительном времени наблюдения и отсутствии ограничений на размножение уцелеет только один из видов”. Впрочем, это дополнение делает очевидным, что “закон” Гаузе-Вольтерра — частный случай хорошо известной ситуации, впервые, по-видимому, описанной О. Бальзаком: “пауки в банке” и потом ставшей крылатым выражением. В русскоязычной литературе одним из первых его (а, возможно, и первым) употребил Ф.М. Достоевский.
“Условия экспериментов, породившие “принцип Гаузе”, не имитируют природную ситуацию, а наоборот, исследуют поведение и судьбу популяций, в экстремальных... ситуациях, которых нет в природе” (Фёдоров, 1987, цит. по 2004: 362).
“Формулировку закона Ома необходимо уточнить следующим образом: “Если использовать тщательно отобранные и безупречно подготовленные исходные материалы, то при наличии некоторого навыка из них можно сконструировать электрическую цепь, для которой измерения отношения тока к напряжению, даже если они производятся в течение ограниченного времени, дают значения, которые после введения соответствующих поправок оказываются равными постоянной величине” (Фишки шутят, 1993: 144).
“Пятидесяти тысяч доходных мест не существует, и вам придется пожирать друг друга, как паукам, посаженным в банку (comme des araignees dans un pot)” О. Бальзак “Отец Горио”.
“Без работы арестанты поели бы друг друга, как пауки в склянке” Ф.М. Достоевский “Записки из мёртвого дома”.
Моделирование континуальности и дискретности
Обитающие совместно виды взаимодействуют друг с другом. Взаимодействия эти могут быть как связаны с переносом энергии (трофическими), так и не связаны. Последние весьма разнообразны и могут быть не менее важными для популяций, чем трофические, хотя на современной стадии своего развития экология изучает прежде всего пути переноса вещества и энергии (цепи питания, конкуренция за ресурс, динамику биогенов и т. д.). Для дальнейшего изложения достаточно констатации факта сложности и многообразия взаимодействий отдельных видов.
Обычно неоднородность биосферы объясняют исключительно неоднородностью абиоты. Подойдём к проблеме со стороны биоты. Может для образования устойчивых в пространстве и времени сообществ достаточно слабых позитивных взаимодействий видов? Положительный ответ на данный вопрос будет означать, что для объяснения наблюдаемого распределения видов в пространстве отнюдь не всегда нужно привлекать абиотические факторы, как это делают континуалисты. Отрицательный же — наоборот, будет свидетельствовать о том, что учитывать влияние среды необходимо для объяснения случаев совпадения пространственного распределения видов.
В природе трудно найти убедительные примеры, поскольку неоднородности биоты наблюдаются на фоне неоднородностей абиоты. Но неоднородную абиотическую среду легко создать в модели. Рассмотрим другую модель (тоже клеточный автомат), которая позволит ответить на поставленный вопрос (Грабовский, 2005).
Представим простейшую систему:
— состоящую всего из двух пар видов А-а и В-b;
— совместно могут обитать только виды разных регистров (вид, обозначенный большой буквой с видом, обозначенным маленькой и наоборот);
— пусть между видами А-а и В-b существует позитивная связь, а между видами А-b и В-а — негативная.
Всё многообразие взаимодействий видов формально выразим в изменении вероятности совместного обнаружения: виды, взаимодействующие позитивно, имеют повышенную вероятность совместного нахождения, взаимодействующие негативно — отрицательную. В модели этот параметр (Ass) варьирует от -1 до +1. При значении Ass = 0 взаимодействие между видами отсутствует, при Ass = -1 или 1 связь функциональна.
На каждом шаге модели каждая особь делится на задаваемое число потомков. Каждая дочерняя особь расселяется в случайно выбранном направлении на случайную дистанцию в пределах задаваемого радиуса и производит попытку занять выбранное место. В зависимости от населения последнего, расселяющаяся особь занимает участок или вымирает согласно матрице вероятностей (как пример в Табл. 1.2 рассмотрен вид а). Если место занято видом А, то особь а займёт его и разделит впоследствии с особью А. Если место пусто, то а займёт его с вероятностью 0,5 + Ass/2. С такой же вероятностью а вытеснит b, если b занимает место совместно с А. В случае, если b является единственным хозяином участка, а вытеснит его с вероятностью 0,5. Если участок занят В, либо В в паре с b, то а становится совладельцем участка с В с вероятностью 0,5 — Ass/2. Судьба вытесненных особей для конечного результата несущественна. Для простоты представим, что все вытесненные особи погибают. В каждом поколении модель сканирует особи в случайном порядке.
Табл. 1.2. Матрица перспектив вида а.
|
Исходное население |
Вероятность события |
Результат |
|
А |
1 |
A/а |
|
-/- |
0,5+Ass/2 |
-/a |
|
А/b |
0,5+Ass/2 |
A/a |
|
-/b |
0,5 |
-/a |
|
В/- |
0,5-Ass/2 |
B/a |
|
В/b |
0,5-Ass/2 |
B/a |
Распределение видов показывается в модели в окне, поделённом на две части. В верхней дано распределение видов А и В, в нижней — а и b.
Модель позволяет задавать два возможных начальных типа распределения (Рис. 1.12) на поверхности тора (тор вместо шара выбран для простоты моделирования, на плоскости же будет проявляться краевой эффект). При одном (слева) получается случайное расположение. При втором (справа) распределение — максимально неслучайное и антагонистичное: одна часть моделируемого пространства заселена видами А и b, другая — В и а, заселённые участки разделены незаселённым пространством. Случай, когда совместно обитают позитивно взаимодействующие виды, не рассматривается. Впрочем, как выяснилось при прогоне модели, влияние начального распределения видов быстро становится несущественным.

Рис. 1.12. Два варианта исходных распределений видов.
На Рис. 1.13 показано, как меняется через 50 поколений исходно случайное (в центре) распределение в зависимости от разных значений (цифры) связи видов. Вверху представлено расположение видов А и В, внизу — а и b. Как и следует ожидать, в отсутствии связи между видами распределение видов независимо. Однако уже при значении параметра связи 0,1 совпадение распределения положительно взаимодействующих видов становится очевидным. Напомним, что такая сила связи свидетельствует, что лишь одна из 10 особей при занятии участка руководствуется предпочтениями в выборе партнера. Дальнейшее увеличение силы связи сначала увеличивает скорость, с которой устанавливаются резкие границы, а затем приводит к возрастанию мозаичности. В пределе (при связи = 1) мозаичность столь высока, что производит впечатление случайного распределения. В действительности же все ячейки заняты либо Аа, либо Вb, но скорость, с которой образуется комплементарное распределение при изначально заданном случайном расположении, столь высока, что обширные однородные участки просто не успевают образоваться. Таким образом, результат не зависит от силы связи видов: при любой образуются участки, занятые той или иной парой видов.
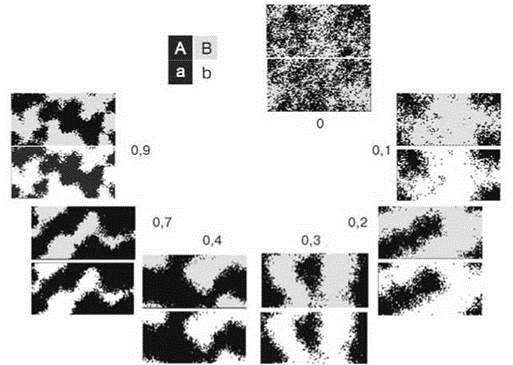
Рис. 1.13. Влияние силы связи видов на их распределение.
Время во всех случаях — 50 поколений, цифры — сила связи.
Большее число потомков и меньший радиус расселения ускоряет формирование однородных участков, меньшее число потомков и больший радиус расселения — замедляют. Т. е. эти особенности биологии влияют лишь на скорость, но не на результат: рано или поздно сообщества, образованные разными парами положительно взаимодействующими видами (т.е. принадлежащие к разным ассоциациям), занимают сплошные области.
Образующиеся между областями границы не просто устойчивы во времени, но более того, их чёткость в ряду поколений только возрастает, при этом граница становится менее извилистой (Рис. 1.14).

Рис. 1.14. Устойчивость границ во времени.
Цифры — число поколений.
Таким образом, для возникновения дискретности совершенно не нужна неоднородность среды. Дискретность может возникать в совершенно однородном пространстве, для её появления достаточно лишь того, чтобы виды взаимодействовали друг с другом (влияли на распределение друг друга), т. е. происходило то, что в природе является обычным. Эта неоднородность образуется в результате самоорганизации и устойчива во времени.
“Ежели у тебя спрошено будет: что полезнее солнце или месяц, ответствуй: месяц, ибо солнце светит днём, когда и без того светло” (Козьма Прутков. Плоды раздумья. Мысли и афоризмы. 51).
Остаётся неясным, с чем же связано то, что при более или менее тщательных поисках обычно удаётся обнаружить, что границы неоднородности биоты часто соответствуют тем или иным неоднородностям среды. Для этого введём в модель неоднородность среды в простейшем виде — как градиент фактора среды, причём при значениях фактора >1 одна из пар имеет конкурентное преимущество, <1 — другая, а при = 0 ни одна, ни другая пара не имеют конкурентного преимущества. При значениях фактора = 0,01 конкурентное преимущество реализуется в 1 случае из 100 при максимальном значении фактора. На графике (Рис. 1.15) это выражено в угле наклона линии фактора: при 0 она параллельна оси х (градиент отсутствует), при 1 — перпендикулярна ей. Сразу же становится очевидным, что даже незначительный градиент существенно влияет на распределение сообществ: граница между ними становится перпендикулярной градиенту среды. При смещении равновесного положения в ту или иную сторону, граница между сообществами движется вслед за ним. Стоит отметить также, что резкость границы не зависит от плавности градиента: при нулевом градиенте она такая же резкая, как и при 1 (конечно, через достаточное число поколений).

Рис. 1.15. Влияние градиента.
Вверху: зависимость резкости границы от значения градиента (цифры), внизу — изменение положения границы вслед за изменением положения нейтрального значения градиента.
Таким образом, хотя градиент среды не является необходимым для возникновения границы при различии конкурентоспособности в разных условиях сообществ, принадлежащих к разным ассоциациям. В этом случае положение границы зависит от градиента: равновесное значение (при котором конкурентоспособность пар равна) является аттрактором. Поскольку абсолютно однородная среда возможна только в компьютерной модели, но не в природе, соответствие градиентов среды границам между ассоциациями видов должно быть весьма обычным. При рассмотрении работ, в которых показывают возможность объяснить распределение видов факторами внешней среды, следует помнить, что любой самый совершенный математический аппарат ищет связи только среди переменных, которые введены в анализ. Первым же этапом любого исследования является ограничение набора рассматриваемых переменных значимыми с точки зрения исследователя или теми, информацию о которых возможно собрать. Поэтому дальнейший анализ устанавливает только то, какие из исследуемых факторов наиболее связаны с распределением видов и не более.
* Часто термин “систематика” используют как синоним термина “таксономия”. Нам кажется более приемлемой точка зрения Дж. Симпсона (1961) и Э. Майра (1971), которые определяли систематику как науку о разнообразии организмов (для которой, кстати, нет какого-либо термина), включая особенности их биологии, т. е. экологию (аутоэкологию) вида. Особенности биологии видов — это такое же их свойство (и таксонов более высоких рангов), как морфология, биохимия и т. д. Таксономию же Э. Майр (1971: 16) определял как “теорию и практику классификации организмов”.
** Континуалисты часто называют её “органицизм” из-за того, что структуралисты видят много сходства в строении организма и экосистем. Мне данный термин не нравится, поскольку он подчеркивает этот, в общем-то, не основной аспект теории, в то же время упуская главное, что отличает её от континуализма — признание наличия внутренней эндогенной пространственно-временной структуры биосферы.
Употребляемое иногда “дискретизм” вызывает обидные ассоциации, поэтому его также вряд ли стоит использовать.
Сам F. Clements говорил о социальном организме и называл свою теорию био-экология, но термин “экология” перестал быть биологическим уже век назад (об этом писал и F. Clements) и вдохнуть в него новую жизнь вряд ли удастся.
С другой стороны, мне высказывали замечание, что термин “структурализм” уже использован этнографом и психологом Клодом Леви-Стросом, но мне кажется, что путаница здесь невозможна.
*** Немецкий математик (1862—1943), член Российской Академии наук (1922) и почётный иностранный член АН СССР (1934).
**** Её суть в том, что пространство и время на моделируемом поле дискретно. Состояние элемента в последующем такте времени полностью определяет его окружение (принцип локальности взаимодействий) и, возможно, его самого в текущем такте времени.