Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Леса, образованные деревьями нетипичных семейств - Причины и скорость образования новых видов и экосистем - Био-экология, или синэкология
Итак, моделирование показывает, что для возникновения дискретности достаточно того, чтобы виды взаимодействовали друг с другом. Детальное изучение биологии отдельных видов всегда выявляет многочисленные примеры взаимодействия изучаемого вида с другими, обитающими вместе с ним. Несомненно, что все эти взаимодействия влияют на эволюцию длительно сосуществующих видов. На признании данного положения и построена парадигма структурализма. Согласно этой теории живущие совместно виды в процессе эволюции адаптируются друг к другу, образуя комплексы, которые мы пока будем называть экосистемами.
Однако процессы эволюции традиционно не рассматривают в экологических исследованиях, поскольку широко распространено мнение об эволюции как медленном плавном процессе накопления морфологических изменений. Имеется в той или иной степени бессознательная тенденция связывать степень морфологических отличий со временем, за которое они происходят: род считают более древним, нежели вид; семейство — более древним, нежели род и т. д. В качестве примера такой точки зрения можно представить Табл. 1.3.
Табл. 1.3. Темпы эволюции или адаптации. По Глазовскому (2004).
|
Уровень |
Время |
|
Семейство Род Вид Морфологические изменения в популяции Функциональные изменения в популяции Индивидуальная адаптация |
1х106 лет nx105 лет nx104 лет nx102 лет nx10 лет минуты - годы |
Цифры, приводимые в подобных таблицах, сильно зависят от анализируемых данных. Так, средний возраст современного вида насекомых — не менее 107 поколений (Жерихин, Расницын, 1980), т. е. соответствует возрасту семейства в приведённой таблице. Известны многочисленные примеры длительного существования видов в неизменном состоянии (латимерия, гаттерия и т. д.). Хрестоматиен пример щитня Triops canacriformis, существующего с позднего триаса до настоящего времени, т. е. около 180 млн. лет. Есть и противоположные примеры.
После вселения из Северной Америки для борьбы с амброзией амброзиевого листоеда Zygogramma suturalis в течение 5—7 поколений образовался новый подвид Z.suturalis volatus Kovalev, 2002, отличающийся не только морфологией, но и способностью к полёту, которой лишены оба американских подвида (Ковалев, 2004).
Пять эндемичных видов гавайских бабочек Hedylepta встречаются исключительно на бананах, завезённых полинезийцами на Гавайские острова около 1000 лет назад (Zimmermann, 1960).
Пять видов рыб рода Haplochromis, эндемичных для африканского оз. Набугабо, образовались не ранее 3720 лет назад (Greenwood, 1965).
Число примеров можно значительно увеличить, но совершенно ясно, что имеется весьма существенный (не менее 6 порядков) разброс скорости видообразования.
“Наличие физических преград, хоть в какой-то мере ограничивающих обмен биотой между территориями со специфическими условиями жизни, становится обязательным условием формирования региональной флоры и фауны” (Мордкович, 2005: 13(8).
“Для обраpования эндемичных видов требуется не только географическая изоляция, но и продолжительное время” (Абдурахманов и др., 2001: 355).
“Close relatives (sister species) have initially vicariant ranges, sympatry is secondary, resulting from later dispersal” (Banarescu, 1992: 823). Э. Майр написал предисловие к этой монографии.
“Опишите процесс видообразования у видов с облигатным бесполым размножением” (Вопрос экзаменатора “на засыпку”).
Классическая схема того, как возникают виды и экосистемы приведена на Рис. 1.16. Согласно этим представлениям, новые виды и экосистемы образуются преимущественно после разделения единого ареала (вида, экосистемы) непреодолимым барьером. Под барьером следует понимать любую преграду, исключающую генетический обмен между популяциями по разные стороны барьера. На схеме исходной биотой является биота А. Возникшие барьеры (барьеры 1 и 2) разделяют ареал этой биоты. В каждом из ареалов возникают самостоятельные биоты, которые постепенно дивергируют.
В случае исчезновения барьера (барьер 1) биоты сливаются в одну. Аналогично развиваются события после проникновения биоты через обычно непроницаемый для неё барьер (барьер 3). Выдающийся американский эволюционист Эрнст Майр (1905—2005) вообще отрицал возможность симпатрического (без изоляции) видообразования за исключением особых случаев, вроде полиплоидного видообразования.
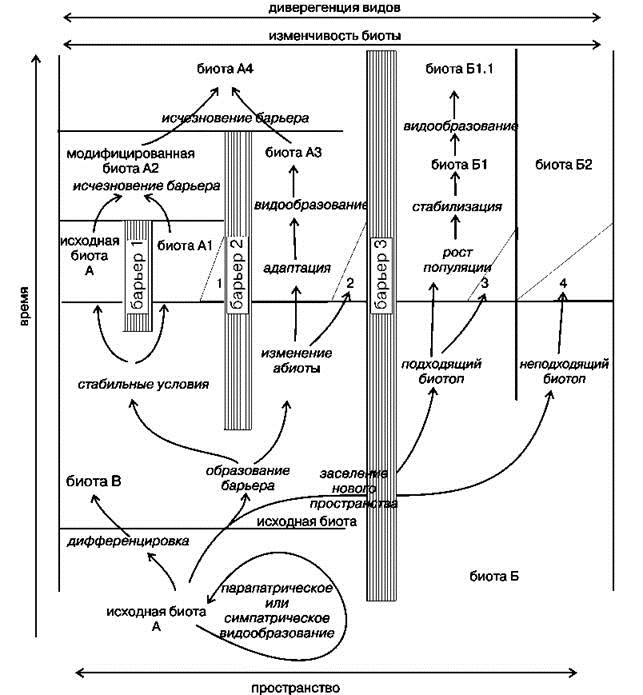
Рис. 1.16. Распространённое представление об образовании видов и биот. По Myers, Giller (1988).
Объяснения в тексте.
Поскольку на его трудах (в русском переводе опубликованы: Майр, 1947, 1968, 1971, 1974 и др.) выросло не одно поколение биологов как за рубежом, так и у нас в стране (да и я в том числе), представление о главенствующей роли изоляции в видообразовании получило широчайшее распространение, и его уже считают одной из прописных истин.
Тем не менее, изучение реальных событий показывает, что представление о необходимости изоляции для образования новых видов, биот и экосистем отнюдь не всегда, а, возможно, и редко соответствуют действительности.
Даже беглый взгляд на распределение видов по Земле показывает, что различные районы населены разным набором видов. При движении по глобальным градиентам среды (от экватора к полюсу, от литорали к абиссали и т. п.) часто можно обнаружить близкородственные виды, сменяющие друг друга вдоль градиента. Несомненно, что одни виды произошли от других, но при этом обычно совершенно невозможно представить себе физические преграды, разделившие гипотетический ареал предкового вида. Особенно сложно такое сделать в случае морских видов, учитывая их широчайшие возможности для распространения благодаря долгоживущим пелагическим личинкам. Число примеров можно значительно увеличить: таковы и ультраабиссальные желоба посреди абиссальных равнин, и глубоководные котловины посреди шельфа — Ледовитый океан, Японское море, и некоторые крупные озёра — Байкал, Ньяса и т. п.). Во всех случаях не возникала преграда. Ареал новых видов оставался связанным с ареалом предковых. Их ареалы в некоторых случаях граничили на протяжении тысяч километров! И, если в отношении тропических лесов имеются данные о том, что в некоторые периоды их площадь резко сокращалась и сплошные массивы распадались на отдельные части, то современная фрагментация бореальных районов моря близка к максимальной за всё время их существования. Представить же барьер, отделяющий малые глубины от больших, ультраабиссальные желоба от абиссальных равнин, вершины гор от их подножий, не стоит и пытаться.
Итак, факты показывают, что образование видов путём изоляции гораздо менее распространено, чем принято считать. Объяснять видообразование изоляцией просто удобнее с педагогической точки зрения. В действительности географическая изоляция не только не является необходимой для эволюции, но даже не стимулирует её.
Конечно, обособление может приводить к постепенной дивергенции разделённых частей ранее единой экосистемы и возникновению из неё дочерних. При этом в каждом из фрагментов ареала предкового вида может образоваться новый вид. Пример тому мы имеем в превращении изначально единой биоты моря Тетис, распавшейся на ряд тепловодных биот. Другой хорошо документированный пример — сравнительно недавнее разделение единой тропической морской центральноамериканской биоты после образования Панамского перешейка примерно 3,5 млн. лет назад. В данном случае по разные стороны перешейка сформировались разные экосистемы и виды, их образующие.
Но даже в таких, казалось бы, убедительных примерах видообразования в результате географической изоляции при детальном анализе оказывается не всё так очевидно. Прежде всего, нет оснований считать, что различия обусловлены только и исключительно самим фактом возникновения Панамского перешейка. Несмотря на то, что его западное и восточное прибрежье находятся в тропиках, абиотические условия их различны. Со стороны Тихого океана из-за влияния пассатов и Перуанского апвеллинга содержание азота к востоку от Панамского перешейка было выше, чем к западу уже 5 млн. лет назад и с тех пор это различие только возрастало (Cannariato, Ravelo, 1997). В результате первичная продукция у тихоокеанского побережья была значительно выше, чем у Атлантического. В то же время абиотические условия более изменчивы: температура воды в Панамском заливе может падать с 27° C до 15°C в течение 24 часов. Высота приливов может достигать 6 м против 0,5 м у Атлантического прибрежья.
Анализ 38 участков ДНК в 9 группах иглокожих, 38 ракообразных, 42 рыб и 26 моллюсков, распространённых по обе стороны перешейка, показал, что 34 были, вероятно, разделены в результате возникновения перешейка, 73 были разделены ранее и в 8 случаях был вероятный контакт после возникновения перешейка (Lessios, 2008). Весьма вероятно, что связь Тихого и Атлантического океана возникала и позже, поскольку даже сейчас территория между восточной Панамой и северо-западной Колумбией находится почти на уровне моря (Graham, 2011)
Каждая из ставших обособленными частей имеет свою дальнейшую историю. В них независимо менялись климат и структура биотопа. После разделения Панамским перешейком части ранее единой биоты взаимодействовали с разными биотами. Распространённость различных сочетаний абиотических условий в изолированных частях может отличаться, что приводит к изменению площади, которую должны занять разные сообщества. Следствием является уменьшение разнообразия одних и увеличение разнообразия других. Эти и другие причины не могли не повлиять на их современный облик и таксономический состав. Так, хотя видовой состав многих таксонов по обе стороны Панамского перешейка весьма сходен, фауны рифообразующих мелководных кораллов Восточной Пацифики и Карибского бассейна совершенно различны (Рис. 1.17)). Около 95% видов кораллов Восточной Пацифики — современные иммигранты из Индо-Вест Пацифики (Robertson et al., 2004). Т. е. фауна кораллов Восточной Пацифики является производной фауны Западной Пацифики, а вовсе не близка к карибской, как можно было бы ожидать, учитывая недавнее возникновение Панамского перешейка. Это свидетельствует о том, что после образования Панамского перешейка мелководные рифообразующие кораллы в Восточной Пацифике вымерли. Размеры современных коралловых рифов тихоокеанского побережья Америки в десятки и сотни раз меньше, чем размеры рифов Атлантического побережья и наиболее древние из них датируют голоценом, а число видов рифообразующих кораллов тут в три раза меньше, чем по другую сторону перешейка (Highsmith, 1980). А ведь с коралловыми рифами связано большое число видов других групп. Поэтому вымирание кораллов неизбежно должно было привести к изменению и даже вымиранию связанных с ними видов и существенному изменению и перестройке не только экосистем самих коралловых рифов, но и рыхлых грунтов.
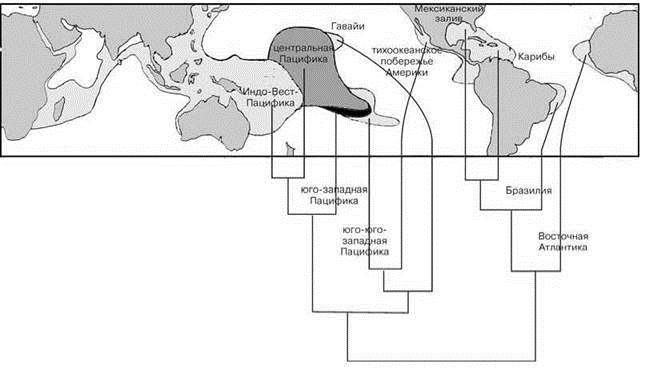
Рис. 1.17. Кладограмма сходства родового состава мелководных рифообразующих кораллов крупных районов Мирового океана.
В качестве меры сходства использован коэффициент различия Браун-Бланке. По Vernon (2000).
А.М. Дьяконов (1955) указывает на наличие тропических видов, обитающих как у тихоокеанского побережья Америки, так и в Индо-Малайском архипелаге, но отсутствующих в Вест-Индии. Причина скорее всего в расселении видов после образования Панамского перешейка.
Луга морских трав весьма обычны в Карибском районе, но отсутствуют в тропической восточной Пацифике (Phillips, Menez, 1988). Поскольку морские травы известны с мела, то нет сомнений, что они обитали в районе Панамского перешейка до его образования и вымерли в тропической восточной Пацифике лишь в самое последнее время. Многие обитатели сообществ морских трав Карибского бассейна имеют близкие виды по другую сторону Панамского перешейка, где они обитают в сообществах рыхлых грунтов. Всё же способность видов менять свою биологию конечна, поэтому часть видов, населявших сообщества морских трав до образования Панамского перешейка, вымерли с его тихоокеанской стороны, как и сами эти травы, а сохранившиеся виды изменились.
Нет в тропических восточных районах Пацифики и брахиопод — типа, известного с палеозоя, т.е. времени, когда ещё не было не только Панамского перешейка, но и разделения Мирового океана на Тихий, Атлантический и Индийский (Зезина, 1973).
К.Н. Несис (1985), сравнивая тропические фауны головоногих моллюсков по обе стороны Панамского перешейка, отмечает, что число бентосных и океанических видов в обоих районах примерно одинаково, тогда как нектобентосных и неритоокеанических гораздо меньше в восточнотихоокеанском районе (5 против 28). Он объясняет это резким интенсифицированием и слиянием Перуанского и Калифорнийского апвеллингов в плейстоцене, приведшим к уничтожению биотопа тропической нижнесублиторальной биоты (внешний шельф — верхняя часть склона, очень узкий в данном районе) из-за постоянного снижения температуры и концентрации кислорода, характерного для апвеллингов.
Данные палеонтологии и изотопный анализ подтверждают это предположение. Они показывают, что на протяжении последних 3—4 млн. лет термоклин на тихоокеанском побережье располагался мелко, позволяя апвеллингам периодически развиваться (Fedorov et al., 2006, Rickaby, Halloran, 2005), данные по составу плиоценовых и плейстоценовых двустворчатых моллюсков Панамского залива свидетельствуют о наличии сезонных апвеллингов (Teranes et al., 1996). .
С другой стороны, число вымерших в Вест-Индии родов моллюсков больше, чем в Восточной Пацифике (Vermeij, 1978), что свидетельствует о том, что и в Вест-Индии условия существования морской биоты претерпевали значительные изменения. Факт массового вымирания тропической фауны Западной Атлантики в это время показан для моллюсков (Stanley, Campbell, 1981).
Итак, таксоцены неритоокеанических и нектобентосных головоногих моллюсков, иглокожих и брахиопод и сообщества морских трав и коралловых рифов (совершенно между собой экологически не связанные) показывают, что после возникновения Панамского перешейка в тропиках на востоке Тихого океана и западе Атлантики произошли существенные изменения условий существования бентоса на шельфе, приведшие к крупномасштабным вымираниям. И климатические колебания, и крупномасштабные вымирания не могли не оказать влияния на эволюцию сохранившихся видов.
Поэтому таксономическое различие биот по разные стороны Панамского перешейка лишь частично можно объяснить самим фактом их географической изоляции. Очевидно, что и в других случаях роль географической изоляции трудно отделить от роли различной истории участков, ставших изолированными.
G.J. Vermeij (1978), сравнивая тропические восточнотихоокеанскую и западноатлантическую биоты, обнаружил, что степень морфологического расхождения зависит от биологии видов. У гастропод, декапод и стоматопод по разные стороны Панамского перешейка она ниже у видов, населяющих рыхлые грунты мелководий, нежели у видов, населяющих каменистые грунты. Все виды тропических американских двустворчатых моллюсков с большим зиянием сзади (т. е. глубоко закапывающиеся) имеют пары близких видов по обе стороны Панамского перешейка. Единственное исключение — Lucinidae. Это легко объяснимо с экологических позиций: для видов данного семейства характерна специализация к хемосимбиотрофии, т. е. они входят в другие ассоциации видов, меньше реагирующие на изменение абиотических условий.
Мало повлияла изоляция и на мелководных морских ежей. К западу от перешейка ныне обитает 20 родов мелководных ежей, а к востоку — 21, по большей части они представлены одним видом, причем в по разные стороны перешейка обитает один и тот же вид (Chesher, 1972; Lessios, 1998).
В то же время имеются многочисленные примеры образования видов без изоляции.
Рассмотрим некоторые районы повышенного эндемизма, а затем попытаемся понять, почему в них шёл интенсивный процесс видообразования, тогда как в других — нет.
Леса, образованные деревьями нетипичных семейств
Хотя жизненная форма дерево или подобная ей имеется во многих семействах покрыто- и голосеменных и даже папортникообразных не все из них образуют леса (т. е. доминируют по обилию над остальными). Одно из таких семейств — сложноцветные (Compositae, или Asteraceae). Это одно из крупнейших семейств покрытосеменных, насчитывающее более 1000 родов и включающее более 20000 видов (почти половину флоры покрытосеменных Земли). Встречаются они везде: от арктических пустынь до тропических лесов, от морских побережий до альпийских снегов. Большинство сложноцветных — травы, реже кустарники, и лишь иногда — деревья. Независимое и неоднократное образование жизненной формы “дерево” возможно у сложноцветных, вероятно, благодаря тому, что у многих видов стебель частично одревесневает, что хорошо знают те, кто пробовал скосить лопух (Arctium sp.): его ствол гораздо прочнее ствола одноразмерной ели — т. е. семейство преадаптировано к образованию деревьев. Но леса, образованные сложноцветными образуются лишь при отсутствии в экосистеме деревьев других таксонов, для которых эта жизненная форма обычна, т. е. в условиях, которые можно охарактеризовать как дефицит деревьев. Такие условия возникли в разных местах (Рис. 1.18). Одно из них — вершины центральноафриканских гор, возраст которых не превышает 1 млн. лет (Рис. 1.19). Местная весьма богатая древесная тропическая флора по каким-то причинам не смогла создать деревья, способные выносить временные заморозки. В результате леса (весьма разреженные) образованы 11 видами подрода Senecio (Dendrosenecio) (род крестовник Senecio обычен и в средней полосе России), на некоторых вершинах до трёх эндемичных видов. Молекулярный анализ показал, что внешнее сходство деревьев разных гор конвергентно. Каждый из видов образовался из местных (Knox, Palmer, 1995), т. е. симпатрично.
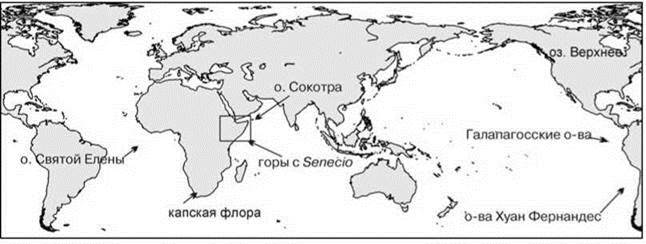
Рис. 1.18. Районы с дефицитом деревьев.
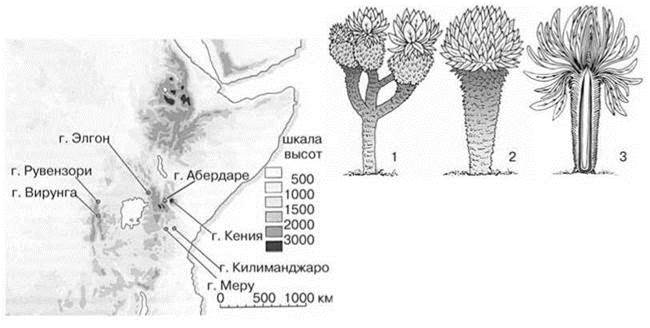
Рис. 1.19. Сложноцветные деревья вершин центральноафриканских гор.
1, 2 — общий вид ветвящейся и неветвящейся формы Senecio, 3 — дерево в разрезе, видны основания старых листьев, создающие “шубу”, предохраняющую ствол от заморозков. На карте — центральноафриканские горы со сложноцветными деревьями рода Senecio. По Cox, Moore (2000).
На острове Святой Елены (место ссылки Наполеона), возникшем в раннем третичном периоде, местная флора была почти полностью самобытной и своеобразной, дефицит деревьев был заполнен сразу пятью видами (Рис. 1.20) сложноцветных. В лесах преобладал Commidendron robustum, было здесь и дерево рода Senecio.
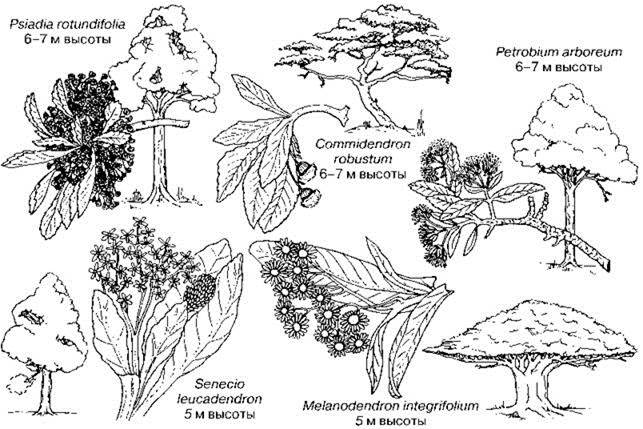
Рис. 1.20. Сложноцветные деревья о-ва Святой Елены. По Cox, Moore (2000).
На Галапагосских островах представители эндемичного рода сложноцветных Scalrsia — деревья высотой более 20 м при диаметре ствола 25—30 см.
На близлежащих островах Хуан Фернандес растёт три древесных рода сложноцветных: Robinsonia, Rhetinodendron и Dendroseris, а также три древесных вида рода марь Chenopodium (Chenopodiaceae), к которому принадлежат обычные в средней полосе огородные сорняки.
На о. Сокотра, расположенном в Аденском заливе, дефицит деревьев заполнен огуречным деревом Dendrosicyos socotranus — представителем семейства тыквенных, близким к дыне (Рис. 1.21).

Рис. 1.21. Adenium obesum и эндемичная тыква Dendrosicyos socotrana (более крупная). Из http://www.zeno.org/Meyers-1905/I/040637a по Schweinfurth G. 1891.
E.C. Pielou считает такие деревья “живыми ископаемыми” “почти несомненно старше, чем острова, которыми ныне ограничен их ареал” (Pielou, 1979: 200). Из текста неясно, на чём основано такое утверждение. Возможно на том, что она просто считает жизненную форму “дерево” древнее жизненной формы “трава” (мнение широко распространённое), но несомненно, что в данном случае это не так. Ареал данных видов ограничен островами не “ныне”, а, наоборот, из приведённых примеров ясно, что жизненная форма “дерево” образуется из местных видов каждый раз, когда экосистема в ней испытывает дефицит.