Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Капская флора - Причины и скорость образования новых видов и экосистем - Био-экология, или синэкология
Пожалуй, один из наиболее ярких примеров эндемизма таксонов разного ранга, вплоть до очень высокого, даёт флора крайнего юга Африки — Капская. В этой флоре только покрытосеменные представлены 6 эндемичными семействами. 19,5% обитающих там родов — эндемы, видовое богатство флоры велико — 8500 видов, из которых 90% — эндемы (Dyer, 1975; Goldbaltt, 1978). Однако это богатство и уникальность не уникальны. На Мадагаскаре из 12 000 видов цветковых растений, 85% видов — эндемы, из 180 семейств — 7 эндемичны, из 1600 родов — 25% эндемичны (Гийоме, 1990). Тем не менее, до недавнего времени это считали достаточным для выделения Капского региона в отдельное царство (Алёхин, 1944; Алёхин и др., 1961; Вальтер, 1968; Тахтаджян, 1970, Good, 1973 и мн. др.). Однако новые данные по биогеографии и экологии заставляют иначе взглянуть на этот феномен. Согласно современным, представлениям, капская флора сформировалась в условиях средиземноморского климата (сухое тёплое лето и влажная холодная зима). Другими такими же регионами являются само Средиземноморье (здесь специфические сообщества имеют местные названия: маквис, маки, маччия, гаррига, фригана, шибляк, томилляры), Калифорния (чаппараль), побережье Чили (маттораль), юго-запад Австралии (мал- ли-скрэб) (Рис. 1.22). В самой Южной Африке сообщества получили название финбош. Вместе данные пять регионов занимают менее 5% поверхности суши, но их флора включает 48250 видов покрытосеменных или 20% земной флоры (Cowling et al., 1996)*. Конечно, до некоторой степени, — это артефакт того, что благодаря прекрасному климату именно здесь размещаются многие научно-исследовательские организации: описана достоверная зависимость между числом видов, известных лишь по гербарным образцам, от расстояния до ближайшего музея с гербарием (Wohlgemuth, 1993). Но это только до некоторой степени объясняет наблюдаемый феномен. Основную причину высокого видового богатства следует искать в процессах формирования экосистем данных районов.
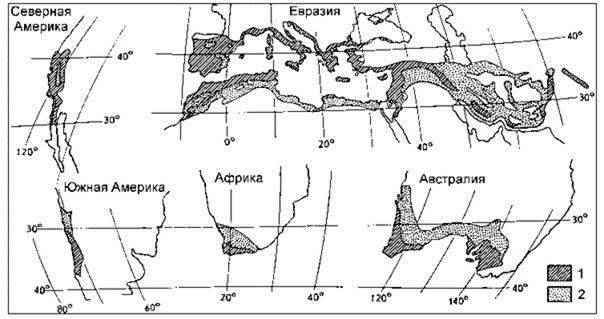
Рис. 1.22. Области средиземноморского климата.
1 — территории со средиземноморским типом климата; 2 — аридные области, преимущественно с зимними дождями. По Г. Вальтер (1968).
В олигоцене все эти регионы были покрыты влажными субтропическими лесами. В начале миоцена сформировался пролив Дрейка. Это привело к росту градиента экватор-полюс и перестройке всей циркуляции атмосферы Земли. Климат этих районов изменился с влажного субтропического на средиземноморский. В первую очередь это привело к существенному уменьшению суммарного количества осадков и их перераспределению в течение года. Старые экосистемы не могли существовать в новых условиях, поэтому возникли совершенно новые, способные существовать в условиях дефицита влаги в наиболее жаркий период года. Растения этих флор имеют специальные адаптации к пожарам, которые в ксеросериях встречаются чаще, чем в мезосериях и тем более гидросериях. Очевидно, что образование таких экосистем из влажных субтропических лесов потребовало резких перестроек, в результате которых сформировались совершенно новые ассоциации видов. В этих новых экосистемах было гораздо больше ассоциаций с обилием жестколистной, в значительной степени ксерофильной кустарниковой растительностью с большим количеством суккулентов. Эти черты выражены в неодинаковой степени в разных местах: в Австралии и Средиземноморье велика роль деревьев, в Южной Африке много суккулентов, тогда как в Австралии их вовсе нет. В Европе, Америке и Австралии образующиеся экосистемы граничили с районами, где обитали виды, преадаптированные к новым условиям. Они послужили основой формировавшихся экосистем, что привело к значительному сходству их флор с соседними. Капская же флора была вынуждена образовываться заново. Своеобразная часть биоты была образована лишь немногими исходными формами, что привело к созданию большого количества эндемичных видов и родов, относящихся к немногим семействам, тогда как остальная биота сравнительно мало отличается от соседних. Среди покрытосеменных в группах, не давших вспышки эндемизма, отличие капской флоры от соседних весьма невелико. Это показало, например, сравнение лилейных разных районов Земли (Рис. 1.23). Зоологи никогда не придавали капскому региону ранга выше подобласти, хотя образование специфических экосистем привело к образованию эндемов и среди животных, особенно много их среди насекомых (жужелица Axinidium, большинство родов трибы жужелиц Creoidosterini, многие чернотелки, долгоносики и др.) (Крыжановский, 1976, 2002). Самостоятельным царством капский регион можно считать только при чисто формальном подходе, который опирается на арифметический подсчёт эндемичных таксонов и игнорирует биологический смысл выделяемых биогеографических регионов. Нет веских биологических причин рассматривать Капскую флору как самостоятельное царство. Она лишь интересная провинция. Стремление придать биологическое содержание биогеографическим выделам и их рангу получает в последнее время всё большее распространение. В частности, понизить ранг Капского флористического региона до провинции предлагают уже многие (Cowiling et al., 1996; Разумовский, 1999; Cox, Moore, 2000). Обсуждение ранга Капского региона см. также Главы 3 и 4.
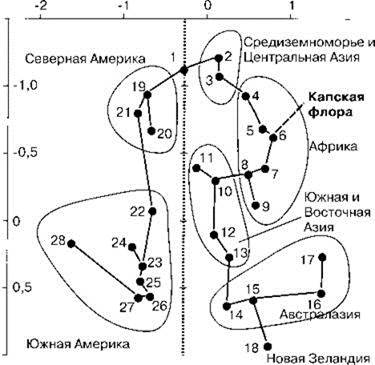
Рис. 1.23. Сходство Liliiflorae разных регионов.
1 — север Голарктики; 2 — Средиземноморье; 3 — Средний Восток и Центральная Азия; 4 — Сахаро-арабский регион; 5 — Намиби-Кару; 6 — мыс Доброй Надежды; 7 — присахарский регион, восток и юг Центральной Африки (преимущественно безлесные ландшафты); 8 — тропический дождевой лес Западной Африки; 9 — Мадагаскар; 10 — Индия; 11 — восточный Китай; 12 — Юго-Восточная Азия; 13 — Малайзия; 14 — Папуа (Новая Гвинея); 15 — Северная и Центральная Австралия; 16 — юго-запад Австралии; 17 — Центральная Австралия; 19 — Скалистые горы; 20 — восток Северной Америки; 21 — юго-запад Северной Америки; 22 — Центральная Америка; 23 — восток Бразилии; 24 — аргентинская пампа; 25 — Амазония; 26 — Гвианское нагорье на севере Южной Америки; 28 — Патагония. По осям значения, полученные методом многомерного шкалирования. По Conran (1995).
“Своеобразие флоры Капской провинции обусловлено распространением следующих семейств и таксонов, встречающихся только на полуострове Кейп или имеющих здесь свой основной ареал. Семейства: Proteaceae, Restionaceae, Penaeaceae, Bruniaceae, Geissolomaceae, Stilbaceae, Retziaceae, Grubbiaceae.
Трибы: Diosmeae среди Rutaceae.
Роды: Erica (почти 600 видов), Cliffortia (розоцветные, со 108 видами), Muraltia (истодовые, 115 видов), Metalasia и Stoebe (сложноцветные), Roridula (2 вида, семейство Byblidaceae, к которому относятся еще только 2 австралийских вида Byblis). Все протейные, помимо вида Brabejum stellatifolium, относятся к подсемейству Persoonioideae. Семейство богато представлено и в Австралии, но другими родами. В Африке широко распространен самый примитивный род Faurea. Из 100 видов Protea только немногие встречаются за пределами Капской провинции.
Из семейства Restionaceae один только род Restio представлен в Южной Африке 117 видами (1 на Мадагаскаре, 27 в Австралии)” (Вальтер, 1968, цит. по: 1972: 157).