Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Метод индивидуальных ареалов - Методы выделения регионов - Методы районирования - Общая биогеография
Неоднородность биосферы выявляют, изучая индивидуальные ареалы. Ареалы можно группировать, оценивая их сходство на глаз (типизация ареалов), изучать распределение границ ареалов (строя биотические разрезы) или определяя структуру фауны.
“Типизация <ареалов>заключается в замене реального “кружевного” ареала идеализированным сплошным и в заполнении пробелов в наших знаниях об ареале данного вида сведениями о распространении локально лучше изученных видов с максимально сходным распространением” (Несис, 1985: 53). Фактически при типизации:
1. “На глаз” группируют ареалы видов, очерченные достаточно хорошо, т. е. те виды, площадь ареалов которых покрыта сравнительно густо точками находок.
2. К выделенным группам добавляют по сходству “на глаз” виды, распространение которых известно менее точно.
3. Границы ареалов видов наносят на карту и определяют положение областей их сгущения.
Типизацию ареалов широко использует школа морских биологов ЗИН: с его помощью типизировали ареалы брюхоногих моллюсков: рода Neptunea (Голиков, 1963), Buccininae (Голиков, 1980), Oenopotinae морей СССР (Богданов, 1990), Isopoda шельфовых зон холодных и умеренных вод Северного полушария (Кусакин, 1979). Его использовали для районирования пелагиали и бентали Мирового океана по головоногим моллюскам (Несис, 1982, 1985), территории СССР по слепням (Олсуфьев, 1980), районирования Северного Ледовитого океана по полихетам (Жирков, Миронов, 1985).
При построении биотических разрезов анализируют распределение вдоль выбранных осей границ ареалов, общего числа видов или числа центров ареалов. Поведение кривой даёт информацию для проведения границ. Границы проводят по пикам на кривых.
Сергеев (1986) предложил использовать термины мощность и эффективность границы. Мощность — число видов, имеющих границы ареалов в данной области сгущения границ ареалов. Эффективность — отношение числа видов, не пересекающих область сгущения границ ареалов, к числу видов по обе стороны области. Считать надо с двух сторон, т. к. его “прямая и обратная проницаемость” могут быть разными. Например, Енисей: восточная фауна на запад не идет, тогда как фауна западного берега состоит из видов, обитающих на обоих его берегах. Критерий, по которому пик считают достаточным, чтобы быть границей, субъективен. Например, А.И. Кафанов (1991) считает значимыми одни пики и незначимыми — другие, на мой взгляд, не меньшие (Рис. 2.18А). Незначимым он считает и пик на Рис. 2.18В, гораздо более резкий, чем любой из пиков на Рис. 2.18А.
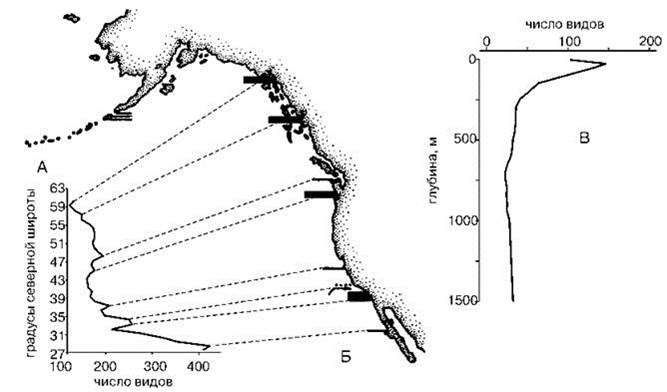
Рис. 2.18. Биотические разрезы.
А — биотический разрез вдоль тихоокеанского побережья Северной Америки; Б — предлагаемое районирование тихоокеанского побережья Северной Америки (линиями соединены пики на кривой суммарного числа видов и проводимые на их основе биогеографические границы); В — биотический разрез у побережья Калифорнии. Скомпановано из нескольких рисунков. По А.И. Кафанову (1991).
J.C. Briggs (1995) проводит только одну границу по 54—55°N, Peden, Wilson (1977) считают, что сгущение границ ареалов приурочено к 50-57°N.
Критерии оценки результатов биотических разрезов разрабатывал В.Н. Семёнов (1982).
Биотические разрезы использовали при изучении вертикальной зональности бентоса (Виноградова, 1958, 1962, 1977), распределения морских ежей (Миронов, 1986 и др.; Парин, Миронов, Несис, 1997), двустворчатых моллюсков Северной Пацифики (Кафанов, 1991), бентоса Южной Америки (Семёнов, 1982).
Изучая структуру биоты определяют долю тех или иных элементов (обычно географических) в произвольно выбранных участках или в отдельных пробах (Рис. 2.19). Следовательно, для применения метода необходимо провести предварительно типизацию арелов видов. А.А. Шорыгин считал достаточным сам факт преобладания видов с каким-либо типом распространения. В случае контакта двух биот это означает, что участок относят к одной из них, если его фауна более, чем на 50% состоит из видов данной биоты. Более корректно установить достоверность преобладания путём оценки достоверности отклонения наблюдаемого на участке соотношения числа видов с различными зоогеографическими характеристиками от такового во всем анализируемом районе. Для этого можно использовать метод выборочных долей по Фишеру (Зайцев, 1984).
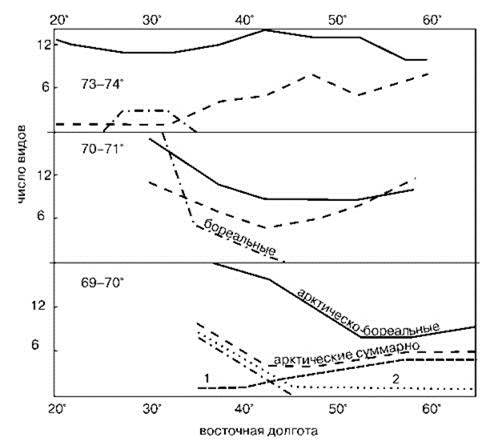
Рис. 2.19. Использование метода структуры биоты.
1 — арктические мелководные; 2 — арктические глубоководные виды. По А.А. Шорыгину (1928).
Недостатки метода. Основной — отсутствие формализованных общепринятых методов проведения границ ареалов (см. Выше раздел “Методы изображения ареалов и проведения их границ”). При проведении границ ареалов отдельных видов всегда наличествует избыток экстра- и интерполяции, а иногда и фантазии. Неудивительно, что границы ареалов отдельных видов, проведённые таким образом, после нанесения их на одну карту образуют причудливый с трудом интерпретируемый рисунок, в лучшем случае дающий так называемые “области сгущения границ ареалов”, занимающие площади, сравнимые с площадями самих биогеографических районов. В результате разрешающая способность метода весьма невелика, он малопригоден для районирования территорий с извилистыми биогеографическими границами и мозаичным размещением районов (например, со сложным рельефом).
Метод не позволяет отличить границы ареала от границ кружева ареала. Априорно предполагают, что лакуны, дающие кружево ареала, гораздо мельче самих ареалов, но есть многочисленные примеры лакун ареала одного вида, существенно превышающие целый ареал другого вида. Рефугиумы считают отсутствующими.
Для увеличения разрешающей способности строят биотические разрезы или анализируют структуру биоты. Области сгущения границ ареалов часто почему-то именуют синператами, хотя синператы по определению, данному автором термина (Кузнецов, 1936), — линии, а не области. Поскольку, не принимая концепцию структурализма невозможно отличить границы ареала от границ кружева ареала, при работе с индивидуальными ареалами считают, что кружево ареалов отсутствует или, что то же самое, не влияет существенно на результаты.
Недостаточно ясно соотношение границы, получаемой с помощью анализа структуры биоты и границ ареалов. Проверка путём проведения районирования этим и другими методами одного и того же района по одним и тем же данным (Жирков, Миронов, 1985) показала хорошее совпадение. В то же время очевидно, что результаты этого метода сильно зависят от того исходного выделения биотических элементов. Структуру биоты изучали для иглокожих Баренцева моря (Шорыгин, 1928), двустворчатых моллюсков дальневосточных морей (Скарлато, 1981); полихет Северного Ледовитого океана (Жирков, Миронов, 1985).
“Совпадение границ ареалов различных видов... позволяет провести... ряд линий общих границ распространения... Подобные линии мы предлагаем назвать синператами” (Кузнецов, 1936: 97).