Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Метод сравнения списков биот произвольно выделенных территорий - Методы выделения регионов - Методы районирования - Общая биогеография
Картирование ареалов не ставит задачу выделения биогеографических районов и анализа их взаимоотношения. Это — задача районирования.
“The biogeographic method does not exist, or there are as many methods as biogeographers” (Dunbar, 1979, цит. по: Longhurst, 1998: 31).
Значение методологии биогеографических исследований едва ли кто-то недооценивает, но к её освещению и разработке проявляется очевидное пренебрежение. Результатом такого отношения является отсутствие в настоящее время общепринятой методики проведения биогеографического членения Земли. Разные авторы делают свои биогеографические построения по-разному, иногда — принципиально различно. К сожалению, при этом по большей части совершенно невозможно понять, как получены предлагаемые схемы, поскольку авторы обычно обходят данный вопрос в статьях, монографиях и учебниках. Такой естественный для научных публикаций раздел как “методы” в них обычно отсутствует или изложен неконструктивно (т. е. недостаточно подробно для повторения другим исследователем). Не так уж редко автор пишет, что он использует один метод, тогда как из дальнейшего текста явствует, что в действительности декларируемым методом он не пользовался или пользовался, но не во всех случаях. Иногда с удивлением понимаешь, что разные части карты построены неодинаковыми методами, иногда в одной книге приведены две совершенно разные схемы без каких-либо разъяснений (Рис. 2.15 6 и 7). Не удивительно, что предлагаемые различными авторами схемы биогеографического районирования порой существенно отличаются. Понятно, что продуктивное сопоставление многочисленных карт, построенных плохо или вовсе никак не описанными методами, по крайней мере, весьма затруднено. Разделы по истории биогеографии содержат много информации о том, кто какие регионы выделял и очень мало о том, какие критерии для этого использовали.
Рис. 2.15 и 2.16. иллюстрируют, насколько картина удручающа в региональном масштабе. На первом изображено районирование наземной растительности Южной Африки, на втором — районирование Баренцева моря.
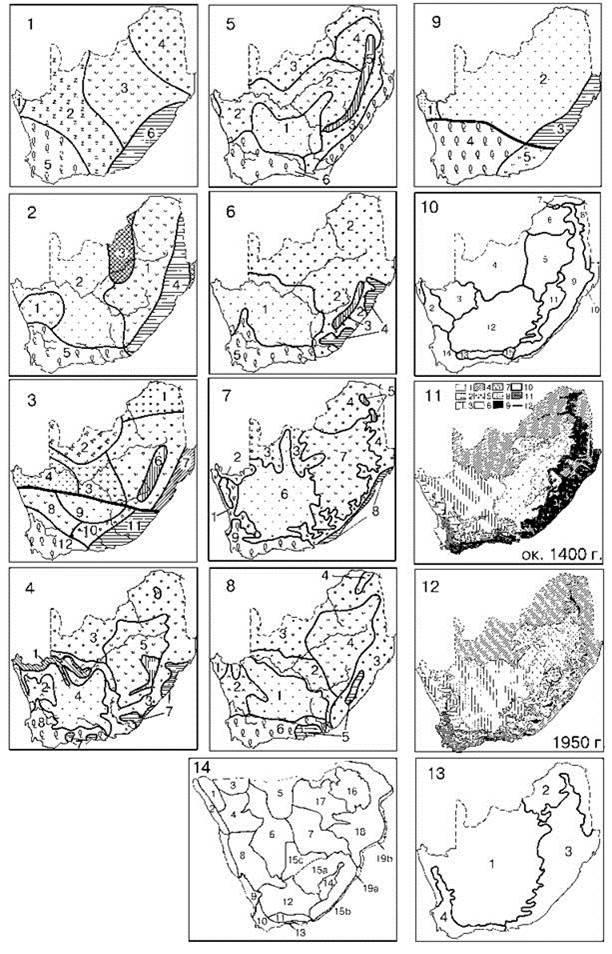
Рис. 2.15. Районирование Южной Африки по растительности.
1— классы формаций в Южной Африке (Dansereau, 1957), 2 — растительные зоны Южной Африки, по И. Гофману (Lesnicky..., 1955), 3 — географические зоны Южной Африки (Страны..., 1973), 4 — растительность Южной Африки, по В.В. Алехину, Л. В. Кудряшову и В. С. Говорухину (1957), 5 — растительность Южной Африки, по В.С. Говорухину (1950), 6 — природные зоны Южной Африки, по карте “Природные зоны мира” (1976), 7 — растительность Южной Африки, по А.И. Зубкову (Страны..., 1973), 8 — растительность Южной Африки, по Т.В. Власовой (Физическая..., 1966), 9 — основные зональные типы растительности Южной Африки, по Т.В. Власовой (Физическая..., 1966), 10 — флористические районы Южной Африки, по Эвансу; 11, 12 — растительности до начала колонизации и после, по Акоксу из Вальтера (1974); 13 — по С.М. Разумовскому (1999, 2011) методом выделения ЦС; 14 — по Hutchinson (1946). Обратите внимание, что рис. 6 и 7 взяты из одной книги.
1: 1 — сухая пустыня; 2 — колючие леса и кустарники; 3 — степь и полупустыня; 4 — саванна; 5 — вечнозеленый лиственный лес; 6 — субтропический дождевой лес. 2: 1 — степи; 2 — пустыни; 3 — муссонные леса; 4 — лавровые леса; 5 — жестколиственные леса. 3: тропический пояс: 1 — зона саванн, редколесий и кустарников (подзона саванн); 2 — зона саванн, редколесий и кустарников (подзона редколесий и кустарников); 3 — зона полупустынь; 4 — зона пустынь; 5 — редколесно-степной тип высотной поясности; б — лесо-луговой тип высотной поясности; 7 — зона тропических лесов (подзона сезонно-влажных лесов); субтропический пояс: 8 — зона полупустынь и пустынь (без расчленения); 9 — зона пустынь; 10 — зона прерий, саванн и кустарников (подзона саванн и кустарников); 11 — зона муссонных смешанных лесов; 12 — зона средиземноморских сухих лесов и кустарников. 4: 1 — леса тропиков и субтропиков, безлистные в сухое время года; 2 — полупустыни и сухие степи; 3 — саванны, редкостойные и саванновые леса тропиков и субтропиков; 4 — пустыни; 5 — степи, прерии, пампа и близкая степям растительность с преобладанием злаков; 6 — растительность высоких плато (горные степи, полупустыни, пустыни и пуны); 7 — вечнозеленые дождевые леса субтропиков, гор тропиков и умеренно теплых областей; 8 — жестколистные вечнозеленые леса и кустарники субтропиков и тропиков. 5: 1 — пустыни; 2 — полупустыни; 3 — саванны; 4 — степи; 5 — растительность гор тропической и субтропической Африки; 6 — вечнозеленые субтропические леса и кустарники. 6: 1 — полупустыни и пустыни умеренной, субтропической и тропической зон; 2 — саванны и саванновые леса; 3 — горные области; 4 — влажные субтропики; 5 — сухие субтропики. 7: 1 — суккулентные пустыни Карру; 2 — суккулентные пустыни Намиб; 3 — опустыненная саванна; 4 — парковая саванна с преобладанием Colophospermum mopane; 5 — высокогорная растительность; 6 — кустарниковые пустыни Карру; 7 — степи Высокого Вельда; 8 — субтропические леса Южной Африки; 9 — вечнозеленые леса и кустарники Калекой области. 8: 1 — пустыни; 2 — сухие степи и полупустыни; 3 — саванны и редкостойные леса и кустарники тропической и субтропической зон; 4 — степи; 5 — влажные вечнозеленые леса субтропической зоны и гор тропиков; 6 — жестколистные вечнозеленые леса и кустарники субтропической зоны и гор тропиков. 9: 1 — тропические береговые пустыни; 2 — тропические внутриматериковые пустыни и полупустыни; 3 — влажные тропические леса; 4 — жестколистные вечнозеленые леса и кустарники; 5 — субтропические степи, полупустыни и пустыни. 10: 1 — Намиб; 2 — Намакваленд; 3 — Бушменленд; 4 — Калахари; 5 — Высокий Вельд; 6 — Бушвельд; 7 — Мопане-Буш; 8 — Низкий Вельд; 9 — Восточный район; 10 — Восточное побережье; 11 — Субальпийский район; 12 — Большое Карру; 13 — Малое Карру; 14 — Капский район; 15 — Аддо-Буш. 11, 12: 1 — пустыня; 2 — собственно Карру с суккулентами; 3 — собственно Карру и карруподобный кустарник; 4 — кустарники бушвелд; 5 — кустарники бушвелд и злаковники; в — низкотравные злаковники: 7 — смешанные злаковники; 8 — высокотравные злаковники; 9 — вечнозеленый дождевой лес; 10 — открытый маквис; 11 — истинный маквис (финбош); 12 — граница аридного Карру. 13: 1 — Калахарская; 2 — Ангольская; 3 — Натальская; 4 — Капская. 14: 1 — Каоковельд, 2 — пустыня Намиб, 3 — Овамболэнд, 4 — Дамаралэнд, 5 — Нгамилэнд, 6 — Сандвельд Калахари, 7 — Торнвельд Калахари, 8 — Большой Намакулэнд, 9 — Малый Намакулэнд, 10 — Южный и Юго-западный Кейп, 11 — Малый Кару, 12 — Кару, 13 — Южный вечнозелёный лес, 14 — горы Дракенсберг, 15а — Высокий грассвельд, 15b — Грассвельд, 15с — Грассвельд Бечуалэнд, 16 — Южное Родезийское плато, 17 — Южный Родезийский вудлэнд, 18 — Восточный Лоулэнд, 19 — литораль (а — восточная, b — восточная тропическая).
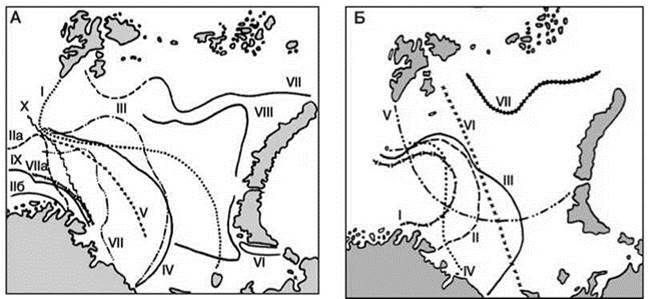
Рис. 2.16. Немногие примеры районирования Баренцева моря по бентосу. По В.Н. Семёнову (1986). Реальное количество границ, проводимых в Баренцевом море на порядок больше.
А: I — границы Арктического и Атлантического бореального субрегионов (подобластей) A. Ortmann (1896); IIа — граница арктической подобласти по Г. Броху, для планктона, IIб — для бентоса, III — граница между арктическим и субарктическим бентосом по К.М. Дерюгину, IV — граница между Арктической областью (Region) и Бореоарктическим переходным районом (Ubergangsgebiete) по Hofsten (1915), V — граница между Арктической и переходной Атлантической областями по E. Hentschel, VI — граница между высоко- и низкоарктической подобластями по В.А. Броцкой и Л.А. Зенкевичу (1939), совпадает с границей А.А. Шорыгина (1928), VII — граница между арктическим и бореальным бентосом по З.А. Филатовой (почти совпадает с соответствующими границами А. Шорыгина, В.А. Броцкой и Л.А. Зенкевича, З. Щедриной), VIIa — средняя граница распространения первой группы бореальных элементов по З.А. Филатовой (1938), VIII — граница между высоко- и низкоарктической подобластями по донным фораминиферам, IX — граница между Арктической и Бореальной областями (VIII и IX — по донным фораминиферам по Л.А. Дигас (1969), X — граница между Арктической и Бореальной областями по В. Брязгину (1973). Б: I — граница максимального и II — граница минимального продвижения арктических видов на запад по А.А. Шорыгину, III — граница арктического и субарктического бентоса по К. Дерюгину (1924), IV — граница арктического и субарктического бентоса по данным 1945—1946 гг. (Черемисина, 1948), V — граница между Атлантической бореальной и Арктической областями по А.Н. Голикову (1980), VI — граница между Арктической и Атлантической бореальной подобластями по О.Г. Кусакину (1971), VII — граница района преобладания арктических видов над остальными группами по А.А. Шорыгину (1928).
В последующих главах приведены разные схемы биогеографического районирования. Их сравнение показывает сколь велико может быть расхождение даже в схемах построенных на одних и тех же данных, но разными методами. Особенно показательны схемы биогеографического районирования суши (Рис. 2.29—2.31 и схемы Главы 3).
“В сущности, А.Л. Тахтаджян первоначально произвольно выделил свои фитохории, основываясь на традиции и интуитивных представлениях, а затем как мог сообщил число эндемичных таксонов, попавших на территории этих субъективных единиц. Это избавляет нас от необходимости подробно рассматривать фактические ошибки его схемы” (Разумовский, 1999: 31).
В целом для биогеографической литературы характерно отсутствие продуктивного сравнительного анализа разных биогеографических схем, недаром P.J. Darlington (1957) считает биогеографические схемы результатом не исследований, не уточнения методов, которыми строят эти схемы, а консенсуса, т. е. постепенного сближения субъективных позиций разных биогеографов. Поскольку различные схемы обычно строят на основе изучения распространения разных таксонов, имеется тенденция все обнаруживаемые при сравнении различия объяснять спецификой таксонов (возрастом, местом возникновения и т. п.), а не различиями в методах районирования. Высказывания с крайне высокой степенью субъективности встречаются даже в работах весьма квалифицированных биогеографов. Так, при обсуждении фаунистических областей P.J. Darlington, один из крупнейших современных биогеографов, пишет: “Heilprin (1887) объединил две северные области, Палеарктику и Неарктику, в Голарктическую область. Здесь они не объединены” (Darlington, 1957, цит. по русскому переводу 1966: 354). Больше к этому вопросу в 518-страничной монографии (в русском переводе) P.J. Darlington не возвращается и свою точку зрения никак не обосновывает. Как результат, подобное приводит к росту субъективности схем, появлению их огромного количества (фактически каждый автор предлагает свою схему, более того, часто даже один и тот же автор предлагает отличающиеся между собой схемы в разных публикациях), при этом обсуждение их отличий в лучшем случае сводится к высказыванию мнений, а то и вовсе отсутствует.
Ниже подробно обсуждены методы биогеографического районирования (или, что то же самое, выделения на местности ЦС и анализ их взаимоотношений). Я надеюсь, что это обсуждение, с одной стороны, поможет разобраться во множестве опубликованных биогеографических схем, а с другой — облегчит выбор адекватного метода исследования.
Любое биогеографическое исследование состоит из двух основных этапов. На первом выделяют сравниваемые объекты, а на втором — проводят их сравнение, по результатам которого строят классификацию, обычно иерархическую. Каждый из этих этапов важен. Очевидно, что некорректная группировка корректно выделенных объектов может загубить любые данные. Возможно, поэтому большая часть работ, рассматривающих методологию биогеографических исследований, посвящена методам группировки районов. С другой стороны, можно с уверенностью сказать, что сколь бы корректно мы не проводили сравнение некорректно выделенных районов, получить адекватные результаты можно только случайно. Поэтому влияние методов выделения районов на конечный результат значительно больше, чем методов их группировки. Но именно выделение районов слишком часто проводят с высокой степенью субъективности. Это приводит к субъективности схем и их множественности. На практике различия схем биогеографического районирования пытаются объяснить спецификой группы, на основе которой строили районирование, о различии же в методах их построения обычно даже и не упоминают. Потому ниже сначала рассмотрены методы выделения биогеографических регионов, а затем — их группировки, но предварительно необходимо сделать ряд замечаний общего характера.
В биогеографии довольно широко используют разнообразный математический аппарат. Особенно это свойственно биогеографическим исследованиям континуалистов. С их точки зрения, биогеографические регионы — условные, договорные, искусственные, т. е. полностью субъективные единицы, удобные для описания континуума биосферы. Математика даёт континуалистам формальные критерии группирования субъективно выделенных участков. В итоге создаётся видимость объективности. Именно видимость, поскольку районы выделяют субъективно, и эта субъективная база биогеографического районирования никуда исчезнуть не может.
С другой стороны отнюдь не всякая особенность биологических объектов, даже подтверждённая обширным корректно обработанным материалом имеет хоть малейший биологический смысл. Например, про одного энтомолога рассказывают, что, промерив нескольких сотен муравейников, он установил, что диаметр муравейника в 3,14 раз больше длины его периметра.
Для структуралистов же биогеографическая схема отражает эндогенную дискретность биосферы, а математический аппарат может использоваться, а может и не использоваться для группировки уже выделенных регионов. В связи с этим надо подчеркнуть два момента.
— Математический аппарат для изучения распределения объектов в географическом пространстве разработан очень слабо. Часто довольно сложно понять, в какой степени результаты математических расчётов отражают объективную ситуацию в природе, а в какой — являются следствием особенностей использованного математического аппарата. Сам факт применения математического аппарата часто используют в качестве аргумента, подтверждающего ту или иную точку зрения (“математически доказано”). Не следует, однако, забывать об ограничениях, накладываемых разными математическими методами на анализируемые данные и о возможностях этих методов. Так, методами ординации нельзя показать, имеются или нет в природе дискретные сообщества. Они для решения этого вопроса просто не пригодны. И наоборот, как отмечает А.И. Кафанов, “иерархические методы кластеризации всегда “подгоняют” структуру любого набора данных под иерархию, независимо от того, каковы реальные отношения между объектами” (Кафанов, 2006: 40).
— Использование математики — не самоцель. Если качественные изменения биосферы в пространстве очевидны на качественном уровне, нет смысла обосновывать это математически. Формальные математические процедуры не могут заменить содержания. Точность науки определяет не степень использования математического аппарата, а точность теорий, лежащих в её основе, т. е. непротиворечивость ее исходных посылок, точность и достоверность следствий из них. В этом математика обычно помочь не в состоянии. Как правило, критерием служит “очевидность на качественном уровне” для автора теории и его последователей. Математика может сказать только, КАК считать, но не может сказать, ЧТО считать.
“Компьютер может выдать вам логические выводы из сделанных вами допущений, однако он не в состоянии оценить правдоподобие самих допущений” (Медоуз и др., 2007: 180).
“Использование алгебраических уравнений не даёт новых идей или информации” (Hengeveld, 1992: 153).
“То, чего я не понимаю без математики, я и с математикой не пойму” М. Фарадей.
“Великий физик Гилберт был очень замкнутым человеком и обычно молчал на заседаниях учёного совета университета, в котором он преподавал. На одном из заседаний, когда решался вопрос о том, чему уделять в новых учебных программах больше времени — математике или иностранным языкам, он не выдержал и произнес речь: “Математика — это язык” — сказал он. (Физики шутят, 1993: 142).
В литературе также встречаются требования учитывать при биогеографическом районировании все встреченные на исследуемой территории виды. Возможно, с точки зрения географии и не имеет значения, входит ли вид в естественные ассоциации или нет. Но для целей изучения истории становления и эволюции природных сообществ и их распространения по поверхности Земли информативны только ценофильные виды, прочие являются “шумом”, затушёвывающим картину естественной дискретности. Поэтому при выделении регионов ценофобные виды (включая случайно встреченные на исследуемой территории или синантропные) учитывать нельзя. И наоборот, виды, истреблённые человеком, учитывать следует.
“— Взгляни на этого математика, — сказал логик. — Он замечает, что первые девяносто девять чисел меньше сотни, и отсюда с помощью того, что он называет индукцией, заключает, что любые числа — меньше сотни.
— Физик верит, — сказал математик, что 60 делится на все числа. Он замечает, что 60 делится на 1, 2, 3, 4, 5 и 6. Он проверяет несколько других чисел, например, 10, 20 и 30, взятых, как он говорит, наугад. Так как 60 делится на них, то он считает экспериментальные данные достаточными.
— Да, но взгляни на инженера, — возразил физик. — Инженер подозревает, что все нечётные числа — простые. Во всяком случае, 1 можно рассматривать как простое число, — доказывает он. — Затем идут 3, 5 и 7, все, несомненно, простые. Затем идет 9 — досадный случай; по-видимому, 9 не является простым числом, но 11 и 13, конечно, простые. Возвратимся к 9, — говорит он, — я заключаю, что 9 должно быть ошибкой эксперимента” (Пой, 1957).
Отдельно следует сказать об учёте видов, появившихся в данной местности в результате деятельности человека. Не так уж редки работы, где сравнивают списки биот, в которые включены как исконно обитающие на сравниваемых площадях виды, так и синантропные, обитающие только в жилищах человека, в непосредственной связи с ним или в таких антропогенно созданных стациях, как помойки, свалки, обочины дорог и т. д. Меня всегда удивляло, почему при подобном подходе в список не включают и другие, связанные исключительно с этими же стациями виды, как Homo sapiens, Canis familiaris, Felis catus; почему учитывают несомненно завезённые рудеральные растения, растущие только по свалкам и обочинам, но не учитывают растущие на клумбах, хотя и то и другое — созданные человеком стации. Несомненно, для целей правильного определения надо включать в определители, как местные виды, так и виды, появившиеся в данном месте в результате деятельности человека. И с этой точки зрения включение в список млекопитающих Европы красношеего валлаби (Macropus rufogresius), как это сделано в “The Atlas of the European Mammals” (Mitchell-Jones et al., 1999), популяция которого существует без поддержки человека в Соединённом королевстве несколько десятилетий, вполне оправдано. Но вряд ли целесообразно учитывать этого кенгуру в составе европейских млекопитающих при биогеографическом районировании.
“Вступая на ложный путь с математикой в качестве проводника, вы лишь скорее всего попадёте пальцем в небо, причём это произойдёт так легко и просто, что Вы даже не заметите, и Вас будет трудно вывести из заблуждения. Логика и математика полезны только после того, как найден правильный путь. Чтобы найти его, следует начинать не с логического мышления, а использовать совсем другие способности, а именно наблюдательность, фантазию и воображение; вернее, точно наблюдение, буйную фантазию и острое воображение” (Г. Эдни Юль (1920), один из крупнейших статистиков, цит. по: Лэк, 1957: 14).
“Сложные расчеты лишь слегка уточняют хорошую экспертную оценку” Научная мудрость.
“Самоочевидный. Очевидный для тебя самого и ни для кого другого” (Бирс, 1966: 282).
Несколько особняком стоят случаи, когда инвазионные виды вошли в состав изучаемых ЦС, например, Rapana venosa в Чёрном море. При проведении биогеографического районирования такие виды, коль скоро они стали ценофильными, следует учитывать.
Поскольку границы биогеографических регионов являются границами ЦС, совпадают границы ареалов не всех видов, а только ценофильных. Границы же ареалов ценофобных видов могут совпадать друг с другом и с биогеографическими границами, а могут и не совпадать, что обычно и происходит. Поэтому при проведении биогеографических границ (выделении ЦС) ценофобные виды являются “шумом”. Так как в одну ЦС входят виды разных таксонов, то биогеографические схемы, построенные на основе различных таксонов, в общем совпадают, однако доля ценофобных видов в разных группах неодинакова. И поэтому схемы, построенные на основе разных таксонов без разделения ценофилов и ценофобов, редко совпадают в деталях. Учёт ценофобов размывает картину. Вспомните приведённый выше пример ястребинок. Учёт 52 основных и 531 промежуточных видов и более 5300 подвидов ястребинок (и это цифры только для части Европы!) способен существенно исказить любые самые очевидные закономерности. Положение ухудшает то, что именно среди ценофобов есть большое число плохо (и не всеми специалистами) различимых видов, что может по-разному исказить картину, в зависимости от того, какую систему использовали: таксономиста-дробителя или объединителя.
Отделить ценофильные виды от ценофобных мы можем, только анализируя ЦС. В то же время, очевидно, что ценофильные виды по определению должны составлять основу ассоциаций (ненарушенных), т. е. встречаться часто. Это приводит к тому, что их ареалы известны лучше, т. е. фактически хорологический анализ обычно опирается на ареалы ценофильных видов, независимо от желания автора, но только если ненарушенные ассоциации занимают площадь, существенно большую, чем нарушенные. Такое наблюдается в море и пресной воде, но, по-видимому, уже очень мало где на суше.
Возможно, неосознанной попыткой проводить районирование только по ценофильным видам является стремление отечественных классиков гидробиологии (Е.Ф. Гурьянова, В.И. Зацепин, Л.А. Зенкевич, З.А. Филатова и др.) использовать при проведении районирования данные по количественному обилию видов.
Но не всё так просто. Ценофобные виды — непременный компонент всякой ЦС, поэтому при выяснении истории формирования ЦС ценофобные виды следует учитывать, хотя и особым образом.
Очевидно, что при проведении биологического биогеографического анализа абиотические факторы должны быть исключены из рассмотрения. “При установлении и описании биогеографической принадлежности следует опираться только на форму ареала, а не на экологические признаки, хотя именно они обусловили эту форму” (Беклемишев, 1969: 105). В действительности же абиотические факторы (температура, солёность, влажность, глубина, высота, геоморфология и т. д.) в явном или неявном виде используют почти во всех биогеографических работах. Ниже мы рассмотрим методы, делящие поверхность Земли на основе особенностей биоты, и не будем рассматривать методы, основанные на подразделении поверхности Земли по гетерогенности абиоты, как не относящиеся к предмету биологической биогеографии.
“Районирование, исходящее из анализа распределения абиотических факторов среды, хотя бы и существенно влияющих на распределение жизни и живого, не может считаться собственно биогеографическим районированием” (Кафанов и др., 1980: 49).
Существует два подхода к выделению сравниваемых биогеографических объектов. При одном подходе выделяют территории, для каждой из которых составляют списки их биот. При другом, на мой взгляд единственно биологическом подходе, сначала выделяют биоты, а затем изучают их распространение. В противном случае необходимо доказывать биотическую однородность выделов, чего никогда не делают. Ключевым вопросом является: “что первично” — разграничение районов или разграничение биот?
С позиций структурализма нет противопоставления биотической (по распространению видов) и биоценотической (по распространению ЦС) биогеографий. Результаты и того и другого районирования должны быть не просто сходны, а полностью идентичны, поскольку оба фактически выявляют одну и ту же дискретность, ибо элементарная биота есть население ЦС, которая существует как динамически меняющийся в пространстве и времени комплекс ассоциаций с резкой и чёткой географической границей. По моему опыту, при детальном анализе ареалов получаемая картина, весьма сходна с картиной, которую можно получить при анализе распределения в пространстве экосистем. Например, биогеографическое районирование Баренцева моря, полученное нами анализом ареалов полихет (Жирков, Леонтович, 2012), т. е. исключительно качественных данных, наиболее близко полученному З.А. Филатовой (1957) на основе анализа распределения двустворчатых моллюсков с использованием количественных данных.
С позиций же континуализма результаты анализа биотической и биоценотической биогеографий могут совпасть только случайно.
Фактически вопросы интегрированности и границ ЦС — суть две стороны одной медали: чем более интегрирована система, тем чётче её границы.
“Если зоогеограф опирается на авторитет мнений, это означает, что у него не хватает фактов или что он ошибается” (Дарлингтон, 1966: 32).
Далее методы рассмотрены в порядке возрастания их точности. Для каждого метода кратко описаны его суть, рассказано как использовать его на практике и обсуждены недостатки. Я попытался также проиллюстрировать различные методы конкретными примерами. К сожалению, биогеографических работ, в которых подробно описывали как получены результаты, очень мало. Часто об этом не говорят вовсе. Многие авторы применяют ранее предложенные схемы. В результате нередко получается абсурдная ситуация: ответственность за решение принципиальных вопросов перекладывают на автора, работавшего столь давно, что подобные вопросы тогда и не ставили. Т. е. в современных работах могут использовать районы, выделенные чисто интуитивно и предварительно в начале XIX в. Часто эти древние и провизорные схемы в дальнейшем корректировали, и авторы попадали в порочный круг: после интуитивного или следующего традициям выделения районов, таксонам давали биогеографическую характеристику, а затем по соотношению таксонов с той или иной характеристикой “уточняли” границы между регионами (замечание Б.Г. Иванова, опубликованное В.Н. Семёновым, 1982). Иногда положение ухудшают тем, что такие улучшения предложенных ранее схем делают методами плохо или вовсе несовместимыми с теми, которыми строили улучшаемые схемы.
Фактически в большинстве случаев из-за слишком кратких описаний использованных методов я пытался понять, какой метод использовали, при том по косвенным указаниям. И, конечно, зачастую ошибался. Часть ответственности за эти ошибки я готов разделить с авторами работ.
И последнее замечание. Если автор практически не описывал используемый метод, то возможность критического разбора сводилась к минимуму или была вообще невозможна. Поэтому часто при разборе методов я мог только ссылаться на работы, где данный метод использовали (по моему мнению) и не мог иллюстрировать общие особенности метода конкретными примерами. И наоборот, чем лучше и полнее был описан использованный метод, тем больше было возможностей для анализа, нередко критического. Может быть, это создаёт впечатление предвзятости. Реальность же полностью противоположна: добросовестность авторов позволяет сказать много, излишняя лаконичность, доходящая до скрытности, такой возможности не даёт. Часто при прочтении биогеографических разделов превосходных в остальных отношениях монографий только и остаётся, что недоумённо пожать плечами. Естественно, что критические замечания к таким работам отсутствуют.
Отнюдь не все использованные при биогеографическом районировании методы являются биогеографическими. Так А.И. Кафанов (2006: 23) пишет: “Повторяя Б.К. Штегмана (1936, 1938), ряд авторов (Reinig, 1937, 1950; Voous, 1960; Матвеев, 1971; Jong, 1972, 1974; Варга, 1976) в качестве основных структур зоогеографии рассматривают не региональные единицы зоогеографического районирования, а динамически изменяющиеся во времени и пространстве “типы фауны” или “фаунистические круги” [Faunenkreise] (Lattin, 1957, 1967). Очевидно, что тип фауны вовсе не эквивалентен территории. Для того, чтобы выделить специфические (а тем более эндемичные) для территории таксоны, сначала надо выделить эту территорию. Но Штегман (1938) не предлагает никаких других методов для выделения районов, кроме тех, которые он во многом справедливо критикует. Не удивительно, что на схеме Штегмана показаны переходные территории, простирающиеся на сотни и тысячи километров.
Методы выделения регионов
Эта стадия в любом биогеографическом исследовании наиболее важна. Однако именно она обычно описана наиболее скупо.
Предельно обобщая, можно выделить три метода выделения биогеографических регионов. В первом методе регионы выделяют произовольно, не заботясь ни о совпадении их границ с неоднородностями биоты, ни о их внутренней однородности. Во втором методе неоднородность биосферы выявляют изучая индивидуальные ареалы. В третьем методе неоднородность выявляют, выделяя общие отрезки границ ареалов ценофильных видов.
Метод сравнения списков биот произвольно выделенных территорий
Использование метода. Тем или иным способом оценивают сходство/различие списков таксонов разных территорий, биогеографические границы проводят пропорционально степени сходства.
Недостатки метода: 1. Регионы выделяют произвольно, не заботясь ни о совпадении их границ с неоднородностями биоты, ни о их внутренней однородности. При этом соответствующие слова могут быть произнесены, но по факту биогеографическую однородность выделенных регионов лишь постулируют, но не доказывают (часто над этим даже и не задумываются). Обычно сравнивают более или менее обширные территории, очерченные (с точки зрения биогеографа) искусственно и случайно: государства и их части, моря, территории, очерченные градусной сеткой, физикогеографические регионы, ландшафты. Прежде всего, это накладывает существенное ограничение на разрешающую способность метода: нельзя ни выделить, ни даже заметить биогеографические регионы, меньшие по площади территорий, для которых составлены списки. Необходимость рассмотрения достаточно крупных выделов, например, 50x50 миль (Hagmeier, Stults, 1964) или 1° долготы х 1° широты (Кафанов, 2005) обосновывают существованием биогеографического и экологического масштабов. Некоторые идут ещё дальше. Так, Виноградов (1999), а вслед за ним и Петров (2001, 2004) предлагают различать “пять иерархических уровней биохор и отвечающих им подразделений географической оболочки — геохор:
— микрохоры и мезохоры топологического уровня — биоценозы, входящие в состав морфологических единиц ландшафта размером 10-1—10-2 км2;
— макрохоры — биогеографические (геоботанические) районы, охватывающие территорию конкретных ландшафтов размером 101—102 км2;
— мегахоры — единицы биогеографического (фитогеографического) районирования размером 103—105 км2;
— гигахоры — главнейшие элементы биосферы и географической оболочки: океаны и материки, зональные типы размером более 106 км2” (Петров, 2001: 99—100).
Сходный подход предлагает Huggett (2002: 13—14). Он выделяет (в скобках пояснения и примеры Huggett):
— microhabitats площадью от нескольких квадратных сантиметров до нескольких квадратных метров (листья, почва, дно озёр, песчаные пляжи, осыпи, стены, берега рек и ручьев).
— mesohabitats площадью порядка 104 км2 (находятся под воздействием одного регионального климата, сходных черт геоморфологии и почв, сходным режимом нарушений: листопадные леса, пещеры и реки.
— macrohabitats имеют площадь порядка 106 км2.
— megahabitats имеют площадь более 106 км2 (континенты и вся поверхность Земли).
“Levels of organization are defined not by their size but by organizational characteristics of the things at those levels” (Allen, Hoekstra, 2015: 8).
При этом необоснование этих уровней не основано на свойствах изучаемых объектов. Виноградов (1999), например, обосновывает это возможностью отображения их на картах разного масштаба. Вопрос о естественности и гомогенности этих выделов с биологической точки зрения даже и не ставят. Всё обоснование классификации сводится к голословным утверждениям о специфических чертах объектов данного размера. Не удивительно, что размеры выделов у этих и других (Miller, 1978; Rowe, Sheard, 1981; Bailey, 2009) авторов различаются в сотни и тысячи раз (Рис. 2.17), а диапазон размеров рассматриваемых объектов — в десятки тысяч, при том, что количество выделов остаётся психологически удобным: три—четыре. Субъективность подобного подхода несомненна. С биологической точки зрения он не оставляет возможности для получения выделов, соответствующих каким-либо реально существующим биологическим структурам.
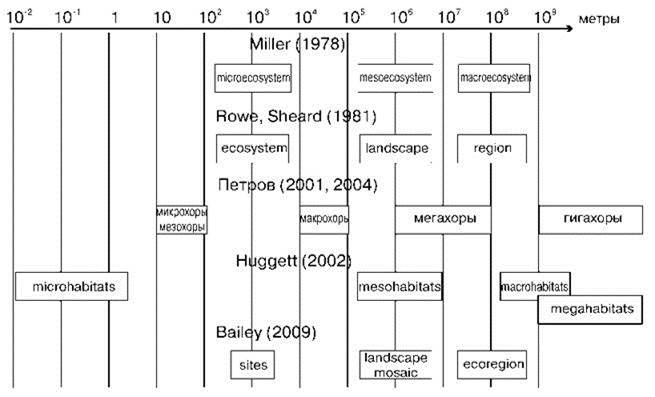
Рис. 2.17. Сопоставление размеров биогеографических выделов, сделанных разными авторами.
Действительно, масштаб — это основное различие экологии и биогеографии, однако размер биогеографических структур может варьировать в весьма широких пределах, и задание каких-либо конкретных значений — принципиально неверно. Например, А.И. Кафанов и В.А. Кудряшов (2000) выделяют Гидротермальную биогеографическую область, хотя размеры отдельных гидротерм в любом направлении куда как меньше 1° долготы х 1° широты, который Кафанов (2005: 61) считает “наименьшей площадью при биотическом районировании в региональной шкале”. При рассмотрении их в таком же масштабе, как и биот умеренных широт Северного полушария, гидротермы будут просто незаметны и их не то что не следовало выделять в отдельную область, но и вообще в какой-либо биогеографический регион самого мелкого ранга.
Сам по себе метод очень груб. Но главное состоит в том, что почти всегда используемые в методе выделы биогеографически неоднородны (относятся к нескольким биогеографическим регионам), и их анализ неизбежно сглаживает биогеографические контрасты, часто создавая обманчивое впечатление наличия континуума. Чем меньше районов сравнивают, тем больше результаты районирования предопределяет изначальное разбиение на районы. При этом насколько проведенные границы соответствуют биогеографическим остаётся неясным (см., например, Голиков, 1963; 1980; Кафанов, 1991; Скарлато, 1981; Petryashov V.V. et al. 2013).
“Важным недостатком метода... является его формальность... районы характеризуются на основании фаунистических списков, часто без согласования с ... особенностями данной местности. Так, например, вопрос о принадлежности восточного Тибета к тому или иному району решается... на основании преобладания тех или иных элементов в общем списке. Между тем... мы можем расчленить общий список на элементы двух совершенно различных районов: высокогорные участки населены тибетской фауной, между тем как в глубоких долинах господствует совсем на неё не похожая китайская фауна” (Штегман, 1938: 2).
Подобный подход критиковал ещё А.И. Толмачев (1931, 1986), противопоставлявший методу выделения биогеографических районов по физико-географическим или административным критериям метод выделения районов по признакам растительности: “следует ли рассматривать как флору всякую... совокупность видов... должны ли мы смотреть на флору как на нечто реальное, т. е. некоторый закономерно сложившийся комплекс видов, или же флора представляет лишь некоторое условное, умозрительное объединение их” (Толмачев, 1931: 113).
Е.И. Матюшкин (1972), например, отмечает, что “трактовка фауны Уссурийского края... прежде всего, как фауны смешанной давно стала тривиальной” (цит. по: Матюшкин, 2005), несмотря на отчётливую разобщённость в пространстве различных фаунистических и флористических элементов. Смена эта весьма резка и происходит в интервале не более одного-двух км. Резкая смена биоты делает неизбежным разделение данного региона на два. “Здесь отнюдь нет того хаотического конгломерата животных разного происхождения, который часто предстаёт перед читателем со страниц популярных очерков о природе Дальнего Востока” (там же, стр. 175). Но именно такой “хаотический конгломерат” мы получим при составлении списка любого региона, выделенного не по признакам биоты, поскольку вероятность совпадения его границ с границами ЦС пренебрежимо мала.
Пожалуй, самым ярким примером, показывающим, насколько такой метод способен исказить реальную картину биотической неоднородности, это деление Голарктики. Во многих схемах биогеографического районирования Голарктику разделяют на Палеарктику и Неарктику. Между тем абсолютно во всех таксонах крупного ранга различие тундры Евразии и Америки исчезающе мало, а различие тайги и даже северных районов зоны широколиственных лесов — незначительно. Различие Палеарктики и Неарктики действительно существует, но оно обусловлено различиями южных районов. Поэтому при сравнении списков биот материков (регионов, выделенных по небиотическому основанию) и получают существенные различия. Но деление Голарктики на Палеарктику и Неарктику совершенно игнорирует отсутствие существенных различий биоты северных районов. Схемы Бобринского (1951), Крыжановского (2002) и других, в которых северные районы обоих материков объединены в один биогеографический регион, а южные — разделены на несколько, гораздо адекватнее отражают сходство и различие биот.
Нередко для увеличения разрешающей способности метода уменьшают площадь рассматриваемого региона. При достаточно большом числе сравниваемых регионов, особенно, если они в той или иной степени изолированы друг от друга получаемые результаты приближаются к получаемым более точными методами, описанными ниже. Примером такой работы может служить Veron et al. (2015), где дан анализ ареалов фауны герматипных кораллов (всего 845 видов) в 150 экорегионах.
Вариантом метода сравнения списка биот являются методы ординации и классификации. Нелишне напомнить, что сравнение таксономических списков разных частей одного биогеографического региона не является биогеографическим исследованием, поскольку между сравниваемыми участками не проходят границы ареалов. Тем не менее, работы такого типа часто используют для доказательства наличия биогеографического континуума.
На фоне основного недостатка метода, остальные недостатки не способны существенно усилить неадекватность результатов. Тем не менее, их следует упомянуть.
2. Все виды обычно считают равноценными (не учитывают их ценотическую роль). Между тем, значительную часть каждой биоты составляют ценофобные виды, распространение которых определяют другие законы, нежели ценофильных. Эти виды, совершенно непригодные для биогеографического районирования, сильно “зашумляют” дискретность биосферы.
С.М. Разумовский указывает (1999: 33): “Все эти обстоятельства (1 и 2 — ИЖ) направлены на одно и то же — на сглаживание флористических различий и на создание смутного представления о флористической непрерывности растительного покрова. Представление же это, раз возникнув, делает психологически излишними любые попытки усовершенствовать породившую его несовершенную методику. Другими словами, мы видим знакомую каждому биологу картину возникновения искусственного континуума при применении точных количественных методов к неточно собранному материалу”.
4. Результаты зависят от полноты списков сравниваемых регионов.