Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Роль пожаров - Теории, объясняющие вымирание мегафауны - Лесные и безлесные экосистемы - Биогеография суши
Поверхность суши часто подразделяют на районы, занятые сообществами, принадлежащими 15—30 типам растительности (тундра, тайга, тропический дождевой лес, саванна и т. п.). Первоначально эти типы выделяли на основе данных, полученных при непосредственном изучении местности, теперь же — преимущественно на основе изучения изображения со спутников. Эти районы часто называют биомами* или типами ландшафтов, хотя эти два термина вовсе не являются синонимами. Каждый из таких районов считают внутри себя достаточно гомогенным, чтобы данные, полученные в одной или немногих точках, можно было экстраполировать на всю площадь данного выдела. Экологи изучают особенности их функционирования (в первую очередь модную сейчас динамику углерода и кислорода). Пытаются связать изучаемые параметры системы с набором переменных, которые считают наиболее значимыми (количество осадков, солнечная радиация, широта и высота места, материнская порода, почва и т. д. и т. п.). Такой подход подразумевает идентичный ответ биоты любого места в ответ на изменения абиотических условий. Однако, поскольку он основан на физиономическом сходстве в конкретный момент времени, он игнорирует процессы, меняющие изучаемые системы. Это делает невозможным предсказания поведения системы на период, за который эти процессы становятся значимыми. К таким процессам относится не только эволюция, при которой для существенных изменений системы обычно требуется минимум несколько десятков тысячелетий, а то и миллионы лет (хотя в отдельных случаях достаточно и нескольких лет). С практической точки зрения столь длительными процессами можно и пренебречь. Но систему меняют и сукцессии, захватывающие интервалы времени от нескольких столетий до нескольких лет, т. е. существенные и для краткосрочных прогнозов.
Лесные и безлесные экосистемы
Биомы, выделяемые в современных биогеографических схемах, многие делят на лесные и безлесные (тундра, степи, пустыни, саванны). Существование безлесных, или травяных экосистем (БЭ) обычно объясняют дефицитом влаги (Вальтер, 1968, 1974, 1975; Мордкович, 1982). Однако палеонтологические данные свидетельствуют об отсутствии БЭ в раннем палеогене (палеоцене и начале эоцена). В то время климаксными ассоциациями повсеместно являлись лесные, несмотря на то, что на Земле, несомненно, и тогда были районы с теми же условиями, в которых ныне существуют БЭ.
Поэтому объяснять возникновение и широкое распространение БЭ только дефицитом влаги — значит слишком упрощать проблему. Гораздо больше согласуется с фактами теория, согласно которой БЭ возникли в районах с дефицитом влаги при воздействии мегафауны (Пучков, 1991, 1992а, 19926, 1993 и др.; Жерихин, 1993, 1994, 2003). Основные положения теории изложены ниже.
Имеющиеся палеонтологические данные свидетельствуют о том, что БЭ формировались неоднократно и независимо, но всегда сходным образом. Сначала появлялись крупные листоядные позвоночные, которые замедляли прохождение сукцессий. В местах, где появление лесных сукцессионных стадий было замедлено из-за микроклимата, в первую очередь, из-за пониженного количества осадков это привело к фактической остановке сукцессии (ретардационный субклимакс). В результате площадь, занятая травяными ассоциациями, увеличивалась. Увеличение ресурса (травы) вызвало появление его потребителя — разнообразной травоядной фауны, ещё более тормозившей прохождение сукцессий. Процесс, таким образом, развивался с положительной обратной связью. В конечном итоге формировались эндогенно стабилизированные травяные ассоциации с высоким уровнем диффузных коадаптационных связей, весьма лабильных и обеспечивающих гибкое регулирование функционирования ассоциаций в условиях изменчивой среды. Эти ассоциации также следует рассматривать как климакс, поскольку не существует внутренних причин для их изменения. В этих ассоциациях развивались специфичные для них блоки копрофагов, травоядных насекомых (прямокрылых, термитов), сапрофагов, некрофагов и др. Доминирующими продуцентами стали травянистые растения, адаптированные к быстрому росту после стравливания. Везде для БЭ характерны более высокое соотношение продукции и биомассы и более плодородные почвы, чем в соседних лесных сообществах. По-видимому, это связано с отсутствием или, по крайней мере, малой ролью деревьев. В этих условиях функция стабилизации отчастипереходит к почве.
“As in Asia and Australasia, the disappearance of large terrestrial tortoises in the New World coincided with the arrival of humans” (Rhodin et al., 2015: 15).
В кайнозое БЭ сформировались на основе злаков, причём независимо, как минимум в трёх местах. Первые по времени, в эоцене, БЭ сформировались в Южной Америке, их эдификаторами были вымершие таксоны крупных неполнозубых и вымершие отряды южноамериканских копытных, а позднее и крупные грызуны, парно- и непарнокопытные, мозоленогие и хоботные. В Северной Америке БЭ начинают формироваться в олигоцене, а в миоцене этот процесс захватывает и Евразию с Африкой. Эдификаторами этих БЭ в Южной и Северной Америках и Евразии с Африкой, являлись разнообразные копытные, к которым позднее присоединились возникшие в Африке хоботные. В Южной Америке БЭ формировали неполнозубые и эндемичные копытные. И, наконец, в неогене БЭ начали формироваться в Австралии, их эдификаторами были травоядные сумчатые и крупные птицы. Везде в состав БЭ входили и разные неродственные таксоны гигантских черепах. Длина карапакса этих черепах достигала 210 см (Megalochelys atlas) — вдвое больше, чем у черепах Галапагосских островов. Уничтожение черепах, совершенно беззащитных перед человеком, владеющим огнём, происходило столь стремительно, что их влияние на растительность остаётся недооценённым и неизученным. Черепахи, вероятно, были первыми представителями мегафауны, которые стали добычей уже ранних гоминид более 3 750 000 лет назад (Rhodin et al., 2015)**. В Южной и Северной Америках и Евразии с Африкой сформировались гигантские животные, способные уничтожать древостой. В Южной Америке это — наземные ленивцы, в Азии — гигантские носороги, в Африке — хоботные. После экспансии хоботных за пределы Африки, гигантские носороги в Азии вымерли, возможно, не выдержав конкуренции, поскольку добывание пищи с разных ярусов с помощью длинной шеи менее эффективно, чем с помощью хобота. Наземные ленивцы, использовавшие передние лапы, напротив, вполне ужились с хоботными и в Северной, и в Южной Америках.
На Мадагаскаре эдификаторами БЭ были, вероятно, гигантские черепахи и крупные наземные лемуры, занимавшие экологическую нишу, близкую к южноамериканским наземным ленивцам и бегемоты. Возможно, им помогали эпиорнисы, однако крупные птицы, судя по Новой Зеландии, БЭ которой (туссоковые злаковники) имеют антропогенное происхождение (McWethy et al., 2010), сами не способны создавать БЭ.
БЭ весьма продуктивны. В современных БЭ около половины первичной продукции потребляют термиты, их биомасса составляет порядка 10 г/м2. Термитники достигают сказочных размеров: максимально до 9 м высотой и 30 м диаметром, обычно, впрочем, гораздо меньше, 1—2 м. Биомасса саранчовых — 0,5—1 г/м2. Крупные травоядные млекопитающие в африканских заповедниках и национальных парках дают среднюю биомассу до 2—3 г/м2. В целом травоядные потребляют 17—94 % первичной продукции (Sinclair, 1975; Риклефс, 1977). Растения, адаптированные к постоянному стравливанию, быстрее наращивают биомассу в сезон дождей (McNaughton, 1984). Отрастающие молодые генеративные побеги имеют лучшие пищевые качества.
“Биомасса... диких животных достигает примерно 12—25 т/га, биомасса скота на фермах — около -7,5, а у местного скота эти значения снижаются до 2,5—3,5 т/га” (Вальтер, 1968: 275). 1 т/га = 1 г/м2.
Процесс формирования травяных биомов и эволюции таксонов-эдификаторов шёл параллельно. Наиболее мощное эдификаторное воздействие на СС оказывают хоботные, благодаря силе, низким потерям от хищников, болезней и сезонных неурядиц (Kingdon, 1979, 1982; Owen-Swith, 1987, 1989; и др.). Хоботные дали вспышку таксономического разнообразия одновременно с началом формирования травяных биомов (Рис. 3.1, 3.2). Они не просто осваивали новую адаптивную зону, они её формировали.
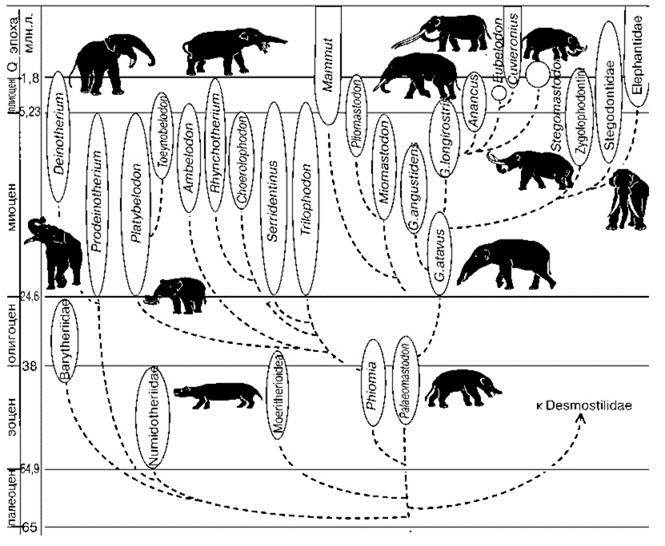
Рис. 3.1. Филогенетическое древо хоботных. По А.К. Агаджаняну (2004).
Deinotherium, Anancus — Африка, Mammut — Голарктика и Африка, Stegodontidae — тропическая Азия, Stegomastodon и Cuvieronius — Южная Америка.
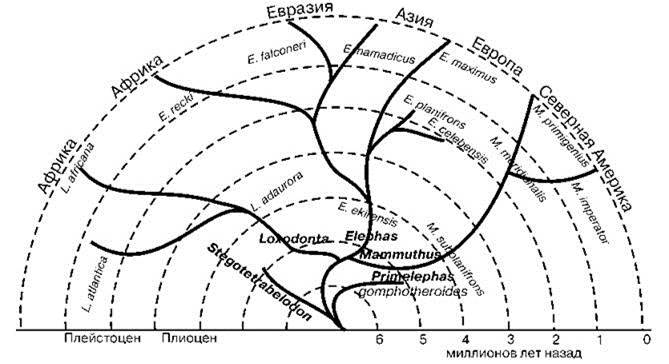
Рис. 3.2. Филогенетическое древо Elephantidae. По Savage, Long (1968).
Слоны уничтожают деревья и кустарники непосредственно, используя то кору, то часть ветвей и листьев, то плоды или корни (Насимович, 1975), из стволов баобабов они выковыривают водянистую сердцевину (Laws, 1970; Насимович, 1975 и др.). Деревца и кусты они выдёргивают, либо ломают; деревья толщиной до 35 см (редко до 70 см) и высотой до 10—12 м выворачивают с корнями; у более крупных обдирают кору. Средства, снижающие вред от копытных (величина, густота ветвей и древостоя, колючки, агрессивные муравьи-симбионты), защищают деревья от слонов не больше, чем от бульдозеров (Wager, 1963; Kingdon, 1979,1982; Вагнер, 1987). Большинство деревьев самцы опрокидывают не для питания, а от избытка сил, либо от ярости, тоски, полового возбуждения; треск крушимой растительности — важный элемент звуковой сигнализации слонов (Wager, 1963; Kingdon, 1979). Слоны активизируют прочие лесоразрушительные силы. Ураганы опрокидывают больше деревьев в разреженном лесу, чем в сомкнутом. Повреждённые деревья чаще гибнут от болезней, насекомых-древоточцев и, особенно, пожаров (Spinage, 1963; Buechner, Dawkins, 1971). Результате на месте леса формируется мозаика из участков с древо- и травостоем различных густоты, высоты и видового состава. Образно выражаясь, такой ландшафт можно назвать “клочковатым”.
О ведущей роли крупных травоядных в возникновении БЭ свидетельствует также то, что после их уничтожения исчезают и БЭ, включая парковые леса и саванны. Они превращаются в сплошные леса или мелколесья (Schenkel, Schenkel-Hulliger, 1969; Laws, 1970; Wilson, Ayerst, 1976). На территории Национального парка в районе водопада Мёрчинсон в Уганде из-за эпидемии сонной болезни в 1912 г. было эвакуировано всё население, это вызвало резкое увеличение поголовья слонов. В результате к 1956 г. общая площадь древостоя сократилась на 55—60% (Buechner, Dakins, 1961). В парках Уганды (Кабалега, Рувензори и др.) солдаты Иди Амина в 1978 г. перебили более 90% толстокожих. Саванны и злаковники за считанные годы заросли акациевыми лесами, что поставило под угрозу выживание большинства копытных (Hatton et al., 1982; Edroma, 1989). Климат запада Кении и прилегающих частей соседних стран за последние 200 лет не менялся. Тем не менее, в начале XIX в. сплошной колючий буш-ньика вытеснил саванны на территории, равной Румынии из-за истребления слонов туземцами, добывавшими бивни (Thornbahn, 1984). Исчезло и большинство других крупных зверей; к концу XIX в. остались только водяные козлы (у реки), малочисленные чёрные носороги, дик-дики, малые куду, да голодные львы, нападавшие на строителей железной дороги. В 1948 г. создали парк Цаво и с середины 50-х вернувшихся слонов хорошо охраняли. Они за несколько лет восстановили мозаику “клочковатого” ландшафта, благодаря чему сюда вернулся весь набор восточно-африканских копытных и хищных (Guggisberg, 1975). Вне парка сохраняется всё та же непролазная ньика (Thornbahn, 1984). В заповеднике Хлухлуве (ЮАР, Наталь) слонов перебили сто лет назад. Выжигания и ограниченные вырубки только тормозили наступление леса и кустарников. Популяции ряда копытных вначале росли, ибо львы, гиеновые собаки и гепарды тоже были уничтожены. Но затем начала сокращаться численность белого и чёрного носорогов, зебр, куду, бушбока, а стенбок (Raphicerus campestris), клипшпрингер (Oreotragus oreotragus) и тростниковый козёл (Redunca arundinum) исчезли совсем. Зато прежде редкая ньяла (Tragelaphusbuxtoni) резко возросла в числе (Owen-Smith, 1989). В долине Чобе (Ботсвана) на протяжении десятилетий в отсутствии слонов буш неуклонно наступал, несмотря на выжигания. Одновременно сокращалось поголовье зебр, жираф, стенбока, гну, канны, лошадиной и прочих антилоп, а также хищников, причём гепард исчез полностью. Возвращение слонов повернуло процесс вспять (Simpson, 1978). В Зимбабве в войну 1972—1980 гг. браконьерство прекратилось и размножившиеся слоны превратили колоссальную площадь лесов и бушей в богатую дичью мозаику из кустарниковой поросли и злаковников (Hallagen, 1981).
Увеличение “клочковатости” ландшафта повышает таксономическое разнообразие и из-за того, что большинство копытных приурочено к экотонам лесных и безлесных участков; одни виды нуждаются и в злаковом, и в веточном корме, другие — используют древостой для укрытия. Некоторые, как например ориби, пасутся на коротких злаках, а прячутся в высокотравье (Kingdon, 1982). Здесь уживаются многочисленные копытные, не встречавшиеся бы вместе в отсутствии слонов. Разнообразие добычи и повышенная мозаичность гарантируют и сосуществование различных хищников. Разнообразие мегафауны поддерживают и слоновьи колодцы в руслах высохших рек (Vesey-FitzGerald, 1960; Kingdon, 1979; Вагнер, Шнейдерова, 1984). Такие колодцы глубиной до 1,5 (Насимович, 1975) и даже до 2,4 м (Haynes, 1987) позволяют жирафам, львам и чёрным носорогам проникать даже в пустыню Намиб (Giraffes..., 1981). Даже ограничивая численность некоторых копытных (кустарникового дукера Silvicapra grimmia, бушбокаTragelaphus scriptus, малого куду Т. imberbis, канны и т. д.) через трофическую конкуренцию и частичное разрушение предпочитаемых биотопов, слоны повышают разнообразие экосистемы в целом (Owen-Swith, 1989). Лишь невозможность миграции слонов по вине человека приводит к полному уничтожению деревьев и превращению клочковатого ландшафта в однообразные злаковники и обеднению мегафауны (Laws, 1970 и др.), поскольку слоны временно покидают местность, где истощается древесный корм (Spinage, 1963). Деградации пастбищ препятствует селективность питания фитофагов (Вагнер, 1987; и др.). Различия в предпочтении потребления растений фитофагами-позвоночными приводит к увеличению мозаичности, что ведёт к росту таксономического разнообразия, устойчивости и продуктивности. Биомасса диких копытных саванны может быть в 4—15 раз больше, чем биомасса коз и овец, выращиваемых на той же территории (Машкин, 2006). При этом домашний скот, в отличие от естественного сообщества крупных травоядных проводит к деградации пастбищ, поэтому с течением времени эта разница возрастает. Пастбища деградируют в пустыни, тогда как продуктивность естественных сообществ с течением времени не меняется.
“Почвы прерий хорошо увлажнены. Поэтому облесение этой области представляется делом вполне реальным” (Вальтер, 1975б: 96).
“Хотя понятия “саванна” и “степь” были введены для обошачения определённых типов растительности, в последнее время они применяются географами скорее для обозначения соответствующих климатических зон... В этом случае, по мнению автора, следовало бы пользоваться чисто климатической терминологией... ботанические термины... должны применяться для характеристики какой-то определённой категории растительного покрова” (Вальтер, 196(8: 276).
“С 1947 по 1949 г. плотность населения оленя и косули в Беловежской пуще возросла примерно на 80%, а количество пострадавших от них деревьев увеличилось почти в 30 раз” (Динесман, 1961: 76).
Крупные травоядные млекопитающие наиболее разнообразны там, где воздействие слонов дополнено тремя другими видами современной мегафауны (Owen-Smith, 1987): носорогами, бегемотами и буйволами. Чёрные носороги “стригут” высокие кусты, включая колючие и огнестойкие, до состояния “щётки” высотой от 1 м (в середине) до 10 см (по краям) (Schenkel, Schenkel-Hulliger, 1969; Kingdon, 1979). Реже носороги выковыривают рогом куст или деревцо (Penny, 1987). Носороги иногда останавливают наступление буша, но неспособны существенно изреживать сформировавшийся древостой (Owen-Smith, 1987).
Прочие виды травоядных млекопитающих предпочитают зелёные части растений: жирафы поедают листья деревьев, антилопы-гну — лишь сочные зелёные листья злаков, зебры — более старые, но ещё зелёные и т. п. (Вальтер, 1968: 275).
Зебры (Equus quagga) готовят в Серенгети (Танзания) пастбище для гну (Connochaetes taurinus), за которыми следуют газели Томпсона (Gazella thompsoni), легко достригающие потравленные злаки, получая доступ к мелким двудольным, важным для переживания ими сухого сезона (Gwynne, Bell, 1968). Газели Гранта (G. granti) пасутся там, где выпас гну создает перевес двудольных над злаками. Водяные козлы (Kobus spp.) зависят от выпаса бегемотов (Hippopotamus amphibius) или буйволов (Bubalus caffer), а ориби — от водяных козлов, конгони (Alcelaphus buselaphus) и других копытных (Kingdon, 1982). На краю Калахари зебры “стригут” траву, а куду (Tragelaphus strep- siceros) — кусты до уровня, приемлемого для дик-дика (Madoqua kirki), обильного лишь там, где много и зебр и куду. В парке Умфолози (ЮАР) с увеличением поголовья белых носорогов (Ceratotherium simum) стало больше гну и зебр, эффективно пасущихся по созданному мегафауной низкотравью (Owen-Smith, 1987). Сам носорог, не справляющийся с травостоем выше 1 м (Kingdon, 1979), в “слоновьей траве” парка Гарамба (Заир) выпасается за бегемотами, слонами и буйволами (White Rhino..., 1977 и др.). У озера Руква (Танзания) та же троица превращает двухметровые прибрежные злаки в зелёный газон, питающий в засуху зебр, канн (Taurotragus oryx), тростниковых козлов (Redunca redunca) и топи (Damaliscus lunatus); бегемоты выдавливают в почве тропы-траншеи, в которых травы зеленеют на протяжении сухого сезона, кормя антилопу пуку (Kobus vardoni) (Vesey-FitzGerald, 1960). В парке Рувензори (Уганда) корневищные злаки, необходимые в засуху бородавочнику (Phacochoerus aethiopicus), поддерживает не огонь, как в других регионах (Kingdon, 1979), а пастьба бегемотов (Lock, 1972).
Исчезновение других крупных животных после истребления слонов и носорогов отмечено и в ряде районов Суматры. Из-за зарастания троп резко снижаются подвижность и шансы на выживание тигров, оленей, малайских медведей (Helarctos malayanus) и красных волков (Cuon alpinus). Даже носорогам без слонов становится намного труднее: уменьшается количество и площадь кормовых участков, увеличиваются расстояния между ними, животное не отдыхает на широкой слоновьей тропе (Strien, 1986).
Как показывают исследования, крупные травоядные сдерживают рост деревьев и кустарников и в других районах, в том числе и за пределами тропиков. Наблюдение за современными степными заповедными участками России и Украины показало, что безлесные ассоциации в отсутствие крупных травоядных не являются климаксными: происходит быстрое их залуговение, забурьянивание, зарастание кустарниками и деревьями.
С другой стороны, даже такие сравнительно (с хоботными и носорогами) небольшие животные, как зубры способны сильно изменить лес. Уже через несколько десятилетий в местах реакклиматизации зубров в сплошных лесах появляются зоогенные поляны с луговой флорой площадью до 5 га, крупные прогалины в лесах, которые соединяются широкими тропами к местам водопоя и кормёжки (Смирнова и др., 1999).
Таким образом, очевидно, что растительный покров в присутствии мегафауны выглядит совершенно иначе, чем в её отсутствие.
Поскольку мегафауна — часть СС, её взаимодействие с растительностью является неотъемлемой частью общего функционирования СС. Поэтому в большинстве случаев правильнее не противопоставлять лесные и безлесные ассоциации, а считать их частями единой СС, имеющей два климакса, лесной и безлесный, занимающие биотопы с разной скоростью сукцессий в пределах ареала единой мозаичной СС (“клочковатого ландшафта”). Сукцессии, начинающиеся всегда с нелесных, обычно травяных сообществ, идут к тому или иному климаксу. Переключение между сукцессионными рядами определяет соотношение скорости сукцессии в направлении древесного климакса и интенсивностью воздействия мегафауны в данном конкретном месте. В значительной степени скорость сукцессии зависит от режима увлажнения: ксеросерии более вероятно заканчиваются безлесным климаксом, а гидросерии — лесным. Микроклимат способствует поддержанию и увеличению контраста: восходящие потоки воздуха над безлесными участками приводят к тому, что дожди, особенно локальные, выпадают только над лесными массивами, причём для получения такого контраста по моим собственным наблюдениям достаточно чередования лесных и безлесных участков в несколько квадратных километров. В конечном итоге в каждой конкретной местности создаётся динамический баланс между лесными и безлесными участками, в результате которого СС и приобретали мозаичный вид. В отечественной литературе принято их трактовать как безлесные экосистемы с “интразональной” растительностью, населяющей преимущественно долины рек (о термине “интразональная растительность” см. ниже). Что характерно для данной “зоны” определяют по преобладающим ассоциациями, а антропогенное воздействие на СС игнорируют (подробнее см. ниже).
“Истребилось всякое существо, которое было на поверхности всей земли; от человека до скота, и гадов, и птиц небесных, — все истребилось с земли, остался только Ной и что было с ним в ковчеге” Ветхий завет. Первая книга Моисеева. Бытие. Глава 7. Стих 23.
“Сделай себе ковчег... так: длина ковчега триста локтей; ширина его пятьдесят локтей, а высота его тридцать локтей” Ветхий завет. Первая книга Моисеева. Бытие. Глава 7. Стихи 14 и 15.
Локоть тогда был около 0,5 м, следовательно размеры ковчега: длина 150, ширина 25 и высота 15 метров.
Наличие двух сосуществующих климаксов не является обязательным. Если скорость сукцессий, ведущих к древесным стадиям, существенно выше нарушающего действия мегафауны, безлесные участки могут и отсутствовать. Это, вероятно, характерно для тропического дождевого леса.
Если скорость сукцессий, ведущих к древесному климаксу, меньше нарушающего действия мегафауны, то формируются СС, имеющие только безлесный климакс, например береговые пустыни районов апвеллингов.
Но, поскольку СС с одним климаксом (лесным или безлесным) свойственны меньшей части поверхности суши, для большей её части должна быть характерна мозаичность, определяемая эдификаторной ролью мегафауны. Ниже безлесные части СС по возможности рассмотрены вместе с лесными.
Теории, объясняющие вымирание мегафауны
Как известно, в конце плейстоцена произошло существенное вымирание мегафауны, включая почти всех крупных травоядных. Существует три основные теории, объясняющие это селективное вымирание в конце плейстоцена (в порядке появления теорий).
1. Всемирный потоп. Согласно Библии произошёл в позднем плейстоцене (6000 лет назад). Знаменитый натуралист Ной, пытался спасти наземную биоту, построив корабль “Ковчег”. В силу чисто физических причин “Ковчег” имел ограниченные размеры, поэтому Ной захватил с собой лишь тех, кто мог поместиться на его корабле. Для мегафауны на нём просто не хватило места.
2. Климатические изменения.
3. Истребление мегафауны человеком. Истребление могло растягиваться на столетия (как на Мадагаскаре) или происходить стремительно (как в Новой Зеландии). Последнее более впечатляюще, поэтому эта теория получила название blitzkrieg или overkill (Blitzkrieg от немецких Blitz — “молния” и Krieg — “война” — немецкая стратегия ведения скоротечной войны. Overkill — английский военный термин, означающий многократное уничтожение, например ядерным оружием).
Кроме этих трёх теорий были предложены ещё несколько, но обоснования их малоубедительны. Например, вымирание мегафауны связывают с необычайно вирулентной болезнью, принесённой людьми с домашними животными, причём следов от этих домашних животных удивительным образом не осталось.
По-видимому, более правильно говорить о совместном действии факторов, и обсуждать стоит степень преобладания одного из них.
“В прошлом богатство крупных млекопитающих было на всех материках (исключая Австралию), но в результате конкуренции (в Южной Америке) или оледенения (в Северном полушарии) везде, кроме Африки, вымерли самые крупные виды” (Дзунино, Дзуллини, 2010: 211). К сожалению, авторы не указали, с кем конкурировали виды мегафауны Южной Америки: только животных весом более 1 т там было более 30, ныне же нет ни одного и в современной фауне их конкурентов нет (подробнее см. раздел Неогея). Для сравнения в Африке ныне только 4 вида животных весом более 1 т, а до появления человека их было на порядок больше (см. ниже). Утверждение об отсутствии крупных млекопитающих в плейстоцене Австралии неверно (см. ниже).
Сторонники климатической причины вымирания мегафауны объясняют исчезновение мегафауны резким изменением климата после окончания ледникового периода: средняя температура Земли выросла на 6°С. Однако вымирание мегафауны и климатические изменения произошли примерно в это время только на территории Голарктики, причем в Евразии на десятки тысячелетий раньше, чем в Северной Америке, тогда как, вымирание в других районах суши происходило в другое, причём разное время. Например в Австралии вымирание мегафауны произошло 50—40 тыс. лет назад, задолго до окончания последнего ледникового периода. Вымирание мегафауны в Южной Америке не сопровождалось вымиранием её в это же время в находившейся в сходных климатических условиях Африке, но было почти синхронным с вымиранием в Северной Америке. На островах (Врангеля, у Калифорнии, Вест-Индии) мегафауна продолжала существовать тысячелетия после её вымирания на соседних материках. В то же время мегафауна исчезала после проникновения на данную территорию человека (Рис. 3.3). Т е. вымирание происходило синхронно не климатическим событиями, а появлению человека в данном районе. Это же возражение справедливо и для теорий, объясняющих вымирание мегафауны перестройками структуры экосистем, вызванных внутренними причинами, например, распространением видов рода Bison, которых, кстати, не было ни в Южной Америке, ни в Австралии.
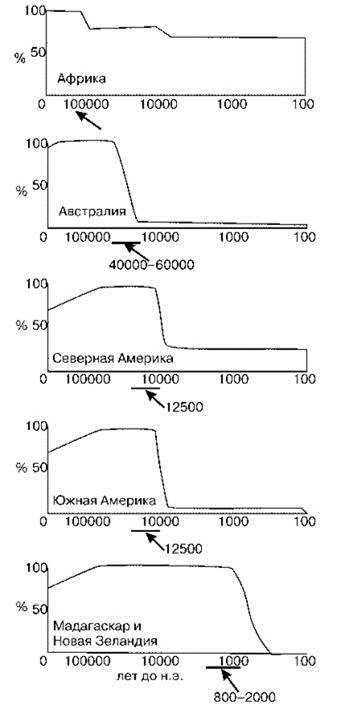
Рис. 3.3. Изменение числа крупных животных в некоторых районах после проникновения туда человека (показано стрелкой). По Brown, Lomolino (1998) и ориг.
Как человек мог привести к исчезновению мегафауны можно рассмотреть на примере Северной Америки, где это вымирание произошло за 1 000 лет.
Проникновение людей в Северную Америку южнее Аляски обычно датируют 13,5—10 тысяч лет назад (Васильев, 2001), хотя имеются данные (нуждающиеся в подтверждении) и о более раннем проникновении. Первые поселенцы принадлежали к культуре кловис, для которой были характерны наконечники копий длиной около 7 см.
Для наконечников кловис характерны желобообразные сколы у основания, облегчающие насаживание наконечника на древко копья (Рис. 3. 4). Эта техника неизвестна из Восточной Сибири, поэтому, вероятно, она была изобретена непосредственно в Америке***. О том, что охотники кловис были первыми людьми на территории Северной Америки (не считая Аляски) свидетельствует (Васильев, 2001):
— Поразительное единообразие наконечников от Атлантики до Тихого океана, что может быть объяснено только быстрым расселением людей по всему континенту, соответствующему экспоненциальному росту их численности.
— Наконечники изготавливали из редкого высококачественного сырья и переносили на расстояния до 500 км, нередко в места, где имелось сырьё не худшего качества.
— Люди кловис редко использовали пещеры и скальные убежища, вероятно, они просто о них не знали.

Рис. 3.4. Наконечник Clovis из нахождения Vail в Maine.
Эти наконечники отличают характерные желобообразные сколы у основания, которые облегчали насаживание наконечника на древко копья. По Hyanes (2002).
Натуральная величина.
Непосредственные находки костей мегафауны на стоянках кловис редки, что служит основанием мнения о том, что кловис были не охотниками на мегафауну, а питались мелкими млекопитающими и растениями. Hyanes (2002) резонно возражает, что находок остатков мелких млекопитающих и растений на тех же стоянках также весьма немного, фактически данные о диете кловис получить из кухонных остатков невозможно. Следует также учесть отсутствие у индейцев вьючных животных, колеса и других транспортных средств, что делает доставку крупной добычи в район стоянки невозможной, вероятнее само племя мигрировало к трупу. Поэтому вполне естественно отсутствие костных остатков в районе стоянки, наоборот, было бы трудно объяснить их появление там. Необходимо помнить, что, поскольку индейцы попали в средние широты Северной Америки с Аляски, очевидно, что в их диете (в отличие от обитателей юга) мясо составляло гораздо более существенную часть, причём объекты охоты были преимущественно крупные, а не мелкие животные. Этому способствовала также возможность сравнительно длительного хранения добычи в условиях холодного климата. Опасность охоты на мегафауну для человека сильно преувеличена, во всяком случае, она не больше, чем опасность для обитателей побережий Северного Ледовитого океана охоты на гораздо более крупных китов и моржей лёгкими гарпунами с маленьких лодок.
Надо также учитывать консервативность поведения мегафауны. Так, киты до сих пор не воспринимают человека как врага, хотя только в 1910—1979 гг. было убито 1 384 085 китов, причём численность некоторых, например синего кита Balaenoptera masculus с примерно 200 000 упала до 1 000 (Бородин, 1996). Вполне вероятно, что мегафауна была истреблена до того, как в её поведении произошли необходимые изменения в связи с появлением нового хищника. Хорошо документированных случаев истребления видов среднего размера (обладающих гораздо большим репродукционным потенциалом) по той же причине, к сожалению, более, чем достаточно.
“If one person in four did all of the hunting, destroying one animal unit (450 kilograms) per week from an animal population on the front averaging 50 animal units per section, he would eliminate 26 percent of the biomass in 1 year” (Martin, 1973: 971).
Последовательность климатических событий, вымирания и расселения человека в Северной Америке дана в Табл. 3.1.
Табл. 3.1. Корреляции изменения климата, предполагаемых событий вымирания и событий в культуре Северной Америки. По Haynes (2002).
|
дата по С14 от настоящего времени |
климат |
временной период или события |
события в фауне |
события в культуре |
|
19,500-16,100 |
очень холодно, сухо |
последний максимум оледенения |
вымираний нет |
археологические находки редки или отсутствуют где бы то ни было на континенте |
|
13,000 |
тепло |
бёллинг |
||
|
12,000 |
аллерёд |
|||
|
11,500 |
холодно |
похолодание в аллерёде |
самые ранние находки кловис, имеющие очень ограниченное распространение |
|
|
11,200 |
тепло |
безымянный период потепления |
некоторое вымирание крупных млекопитающих |
кловис широко распространены |
|
10,900 |
холодно, суше |
резкое начало раннего Дриаса |
кловис становятся локально более разнообразны |
|
|
10,800 |
по прежнему холодно |
вымирание мегафауны завершено, включая мамонтов и мастодонтов |
кловис распространены по всему континенту |
|
|
10,200 |
тепло, более влажно |
ранний Дриас резко заканчивается |
наконечники с желобками больше не делают |
Расчёты (Martin, 1973) показывают, что для быстрого уничтожения мегафауны достаточно соблюдения следующих условий:
1. Прирост численности популяции Homo sapiens в 1,4% в год****. В этом случае удвоение популяции происходило каждые 50 лет и за 1000 лет группа из ста человек даст население в 110 660 583 человек.
2. В процессе движения создавался фронт повышенной численности с плотностью популяции 0,4 человек/кв. км*****, плотность за фронтом была в 10 раз ниже (Рис. 3.5).
3. В год в районе фронта изымалось 20% биомассы мегафауны. Этого достаточно для её полного уничтожения при неселективной охоте за 10 лет. В реальности, вероятно, самок и молодняк были более лёгкой добычей и их убивали более интенсивно, что ускоряло истребление вида.
4. Люди двигались в направлении мест с наибольшей численностью добычи со скоростью примерно 16 км/год. Этой скорости достаточно, чтобы достигнуть Огненной Земли за 1000 лет (Рис. 3.6)******.
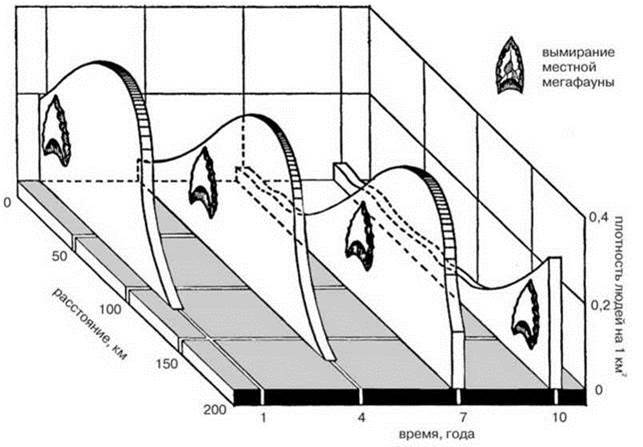
Рис. 3.5. Структура фронта расселения индейцев в Америке.
В каждой точке люди и мегафауна сосуществовали не долее десяти лет. Отсутствие явных археологических следов в этом случае неизбежно. По P.S. Martin (1973).
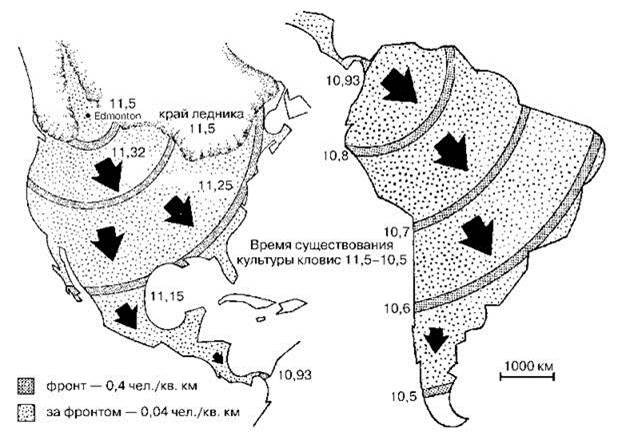
Рис. 3.6. Заселение людьми Северной и Южной Америк. По P.S. Martin (1973).
Все эти допущения вполне правдоподобны. Более того, наблюдения в природе показывают, что популяция вида, попавшего в условия избытка пищи движется именно волной, сходной с распространением пожара в степи. Хорошо документированным примером такой волны является история расселения жука-листоеда Zygogramma suturalis. Этот жук питается исключительно сложноцветным амброзией полынолистной Ambrosia artemisiifolia. Амброзия — американское по происхождению растение и на юге России она — ценофоб. Но, поскольку естественная растительность юга России давно антропогенно нарушена и практически везде заменена ценофобной, амброзия смогла стать злостным сорняком на юге России и Кавказе. Дополнительно массовому развитию и широкому распространению амброзии способствовало и отсутствие естественных врагов. Борьба с амброзией стала необходима не только из-за того, то она стала злостным сорняком, но и из-за того, то её пыльца вызывает аллергическую реакцию у многих людей. Для борьбы с амброзией и ввезли Z. suturalis. Оказавшись в условиях избытка пищи, жук стал быстро размножаться и расселяться, образуя фронт повышенной численности, достигавшей 5000 экз./м2. В тылу фронта амброзия часто полностью отсутствовала. Скорость движения фронта была постоянной (Ковалев, 1994).
Изучение состава изотопов 13C, 15N коллагена неандертальцев (Bocherens et al., 1991) показало, что по доле мяса равными или даже превосходящими волков, что вполне соответствует изложенной выше гипотезе. Весьма маловероятно, что такая диета могла быть результатом собирательства, по-видимому, они были хищниками высшего уровня, облигатными охотниками (Boyle, 2007).
Surovell et al. (2005), Surovell, Waguespack (2008) показали, что интесивность использования хоботных кловис наивысшая из известных в доисторическом мире.
Уничтожение мегафауны снижало эффективность использования растительных ресурсов СС. В результате в СС происходило накопление неиспользованных растительных остатков, что существенно увеличивало интенсивность и частоту пожаров. Появились и антропогенные пожары, как случайные, так и специальные. Пожары, в свою очередь также воздействовали на растительность. Воздействие это было сходным с воздействием мегафауны в том, что преимущество получали травянистые растения. Но, в отличие от воздействия мегафауны, воздействие пожаров было гораздо менее селективным и способствовало устранению участков древесной растительности, т. е. делало ландшафт более гомогенным. К тому же мегафауна распространяла семена многих растений. В результате на месте “клочковатого ландшафта”, с разнообразной биотой, сформировались огромные пространства, покрытые травянистой растительностью и населённые преимущественно одним видом — бизоном. Замена “клочковатых ландшафтов” отдельными зонами прерий и лесов привело к тому, что виды, экологически связанные с опушками, резко сократили свои ареалы. До перестройки они были распространены широко, после неё их ареалы сократились до узкой пограничной полосы между прериями и лесами. Это сопровождалось снижением их численности и в конечном итоге привело к вымиранию некоторых. Селективное истребление мегафауны привело к её стремительному вымиранию после появления в регионе человека (Рис. 3.3, 3.7).
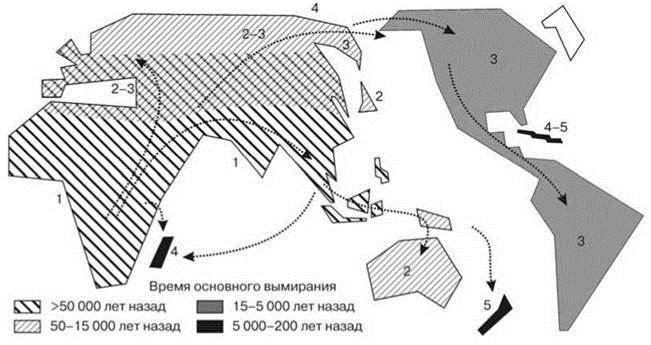
Рис. 3.7. Карта последовательности периодов вымирания на основных массивах суши.
Стрелки показывают направление расселения человека. По Martin (2005), слегка видоизменено.
Скорость истребления мегафауны была столь высока, что её весьма сложно зафиксировать, изучая археологические и палеонтологические данные. Время существования культуры кловис — порядка одной тысячи лет (11500—10500 лет назад) — хорошо согласуется с приведёнными выше оценками. Этого времени оказалось недостаточно для перестройки биоты Северной Америки. Произошёл коллапс, вслед за которым исчезла и культура кловис. Ей на смену пришли разнообразные региональные культуры, общим для которых было отсутствие желобков на наконечниках (Васильев, 2001). “Дикий запад”, который застали европейцы, в действительности был вовсе не в естественном состоянии, а охотничьей монокультурой, созданной индейцами приблизительно 10—8 тысяч лет назад. Seton (1929) полагал, что до прихода европейцев в прериях паслось 60 миллионов бизонов, примерно по столько же вилорогов Antilocapra americana и белохвостых оленей Odocoileus virginianus и по 10 миллионов чернохвостых оленей Odocoileushemionus и лосей, т. е. всего около 200 миллионов голов. При замене этих стад на современные стада коров фактически произошла только замена культивируемых видов, но не способа получения мяса: и индейцы и современные американцы производят мясо, разводя крупный рогатый скот*******.
Охота на мегафауну и, особенно на хоботных и сопоставимых с ними по размеру животных — оптимальная стратегия********. Hyanes (2002) указывает, что один мамонт даёт охотнику столько же энергии, сколько четыре бизона. При этом убить четырёх бизонов сложнее и опаснее, чем одного мамонта. Для убийства афиканского слона достаточно одного пигмея, который протыкает ему копьём кишечник, обычно в районе анального отверстия и ждёт, когда слон умрёт от перитонита. Поэтому, вместо того, чтобы каждые три дня охотится на бизона, охотник, весьма вероятно, предпочтёт раз в 12 дней убить мамонта, которого к тому же выследить гораздо легче*********. Весьма вероятно, что даже при очень низкой плотности хоботных они были добычей высшего приоритета, поэтому бытующее в экологии с математическим уклоном представление о том, что при очень низкой частоте встреч одного вида добычи, охотник переходит на другой, основано на представлении, что для охотник оценивает добычу в удобно подсчитываемых калориях. Однако различие между теорией и практикой на практике гораздо больше, чем в теории. Вполне возможно, что убийство мамонта (или мастодонта) могло входить в какие-нибудь культурные традиции, например при инициации, как это имеет место у некоторых африканских племён или в некоторых средиземноморских странах (коррида) в отношении других опасных животных. В этом случае ценность добычи, а, следовательно, вероятность окончательного её истребления существенно возрастала. Даже в странах, где таких традиций нет, убийство крупной добычи повышает “самцовость” охотника: достаточно вспомнить традицию развешивания трофеев по стенам или стремление убить крупную опасную добычу даже в условиях, когда она неспособна оказывать сопротивление, пример такого поведения дают российские высокопоставленные чиновники, охотящиеся на крупных животных, невзирая на какие-либо законы и здравый смысл.
Освоение эффективных способов охоты позволяло уничтожать зачастую гораздо больше животных, чем было необходимо. Протяженность земляных валов, построенных для загона стад достигала 800 км (Данилкин, 2009). Для пропитания 15 000 человек, обитавших на юге Русской равнины требовалось 10 500 т мяса, что примерно соответствует более 1 000 мамонтов, 10 000 — 30 000 бизонов, 60 000 — 80 000 лошадей или 120 000 северных оленей (Верещагин, 1971; Верещагин, Барышников, 1985).
Роль пожаров
Отдельно следует обсудить роль пожаров в поддержании безлесности (роль пожаров в преобразовании СС рассмотрена ниже, при рассмотрении растительности субтропиков). Вальтер (1968), описывая растительность тропической зоны, неоднократно подчёркивает, что тропические саванны как травяные экосистемы, лишённые деревьев, в настоящее время поддерживает воздействие человека, прежде всего выжигание накапливающейся сухой травы. Более того, исследователи, занимающиеся динамикой развития безлесных экосистем и их историей подчёркивают, что само развитие саванн, как и тропических пустынь связано с человеком (Cloudsley-Thompson, 1969; Сахара, 1990). То же, по-видимому, справедливо и для умеренных безлесных районов. Новозеландские злаковники сменили естественные леса в результате антропогенных пожаров (McWethy et al, 2010; Bowman, Haberle, 2010). Существенные изменения растительности Австралии также были вызваны антропогенными пожарами (Rule et al., 2012;McGlone, 2012). Высокотравные прерии поддерживались в безлесном состоянии ежегодными палами, производимыми индейцами. Когда индейцев сменили американские поселенцы, прерии стали быстро зарастать лесом. То же справедливо и для полупустынных районов Дикого Запада, которые в отсутствие палов зарастают мескитовым деревом, кактусами и разными низкорослыми деревьями.
“Выжигание <саванновых лесов> приводит обычно к образованию травянистых пространств характера степей, которые также деградируют всё больше и больше. Под конец может получиться нечто вроде пустыни” (Алёхин, 1944: 255).
“Важную роль в североамериканской прерии играл и такой фактор, как широко практиковавшиеся раньше индейцами выжигания сухого травостоя” (Вальтер, 1975б: 69).
“Ежегодно треть территории Мадагаскара выгорает от того, что жители в сухой сезон поджигают траву, чтобы обеспечит скот зелёным кормом” (Джолли, Джолли, 1990: 254).
Частота пожаров резко возросла после массового вымирания мегафауны, сопровождавшего становление Homo. До этого сбалансированность экосистем была гораздо выше, поэтому накопления высохшей растительности не принимало таких масштабов или вовсе не происходило. Сведение леса началось более 50 000 лет назад, когда человек овладел огнем. Хотя пожары время от времени случаются и по естественным причинам, поджоги, намеренно устраиваемые человеком, оказывали на растительность гораздо большее воздействие. Прежде всего, это объясняется тем, что искусственный поджог производили на одном и том же месте значительно чаще, чем случались естественные пожары. Естественные пожары возникают гораздо реже и не производят больших разрушений в лесных областях (причина опустошительных пожаров в лесах со средиземноморскими климатом рассмотрена ниже). Лишь после того, как под влиянием деятельности человека СС были разрушены настолько, что начали накапливаться неиспользуемые растительные остатки, интенсивность естественных пожаров резко возросла. В районах с преобладанием безлесных ассоциаций путём многократного выжигания в сочетании с пастьбой интенсивность и длительность действия пожаров настолько усилились, что естественная мозаичность исчезла полностью.
Человек способствует образованию пустынь не только путем выжигания и перевыпаса. По меньшей мере, половина всего леса, вырубаемого в мире каждый день, идет на топливо для приготовления пищи и обогрева жилья. В городах Западной Африки нагреть кастрюлю порой обходится дороже, чем наполнить ее, и поэтому сухой навоз всегда служит топливом, вместо того чтобы служить удобрением для почвы.
Уничтожение леса, кустарника и травяного покрова приводит к возрастанию эрозии: эоловой, если осадков мало и водной, если их много. Поскольку антропогенное воздействие не прекращается, постепенно местность превращается в полупустыню и пустыню. Очень наглядно воздействие человека на растительность засушливых мест на примере пустынь Средней Азии описывает Г. Вальтер (1975: 220 и далее). Несомненно, что накопление сухих растительных остатков, способствующих развитию и естественных пожаров, в значительной степени является следствием разрушения естественных СС, в первую очередь в результате уничтожения мегафауны. Поэтому, хотя пожары и влияют сильнее на древесную растительность, чем на травянистую, роль естественных пожаров в поддержании безлесности, как и в формировании других типов растительности, по-видимому, сильно преувеличена.
Антропогенные пожары могли быть и одной из основных причин вымирания мегафауны, как это произошло в Австралии. Здесь интенсивность пожаров резко возросла после проникновения человека в Австралию. Уцелевшие виды резко изменили свою диету (подробнее см. ниже в характеристике биоты Австралии).
Вымирание мегафауны должно было неизбежно привести к существенным перестройкам биот, заключающимся в утрате ими мозаичности.
“Do global ecosystems function differently now that megaherbivores are gone and human-lit fires are common? New results strongly suggest that they do” (McGlone, 2012: 1453).
В зависимости от того, какая из трёх приведённых выше причин вымирания мегафауны является основной, принципиально меняется та биогеографическая картина мира, которую следует считать естественной, т. е. существующей помимо человека. Если мы примем любую из первых двух концепций, объясняющих это вымирание естественными причинами, то современная биогеографическая картина является естественной. Если же мы примем третью, согласно которой было не вымирание, а истребление, то современная биогеографическая картина нуждается в существенной коррекции, чтобы убрать из неё искажающее влияние человека. Поскольку в данном случае нас интересует биогеография, а не экономическая география, по возможности следует описать биогеографическую картину так, как она бы выглядела без влияния человека.
Любопытно, что уничтожение мегафауны человеком легко было признано в случае Новой Зеландии и Австралии (мегафауну уничтожили аборигены). Но истребление мегафауны на территории Европы признают с трудом, поскольку придётся принять вину на своих предков. Сходная ситуация в Америке, где вину придётся возложить на индейцев, что не политкорректно.
* Автор термина “биом” F. Clements предложил его для обозначения совсем другого явления. В изначальном смысле “the term biome has been applied only to those communities in which studies have established something of the processes of development and the character of the final stage or climax” (Clements, Shelford, 1939: v—vi). Т е., в отличие от “биоценоза” Mobius, biome — это функциональное единство, а не просто статистический артефакт. Фактически в изначальном смысле термин “биом” близок по смыслу к принятому в данной книге термину С.М. Разумовского “ЦС” — ценотическая система. Позднее, термин “биом” стали использовать и континуалисты, а также географы, и в их использовании его смысл становился всё более расплывчатым, а требование функционального единства и сукцессионного развития было утрачено. В настоящее время под биомом всё чаще подразумевают уже не динамически развивающуюся экосистему, а физиономически сходные ландшафты какого-либо региона. Сукцессионные взаимоотношения ассоциаций при этом игнорируют. Физиономическое сходство авторы конкретной системы биомов, оценивают по признакам, которые они считает существенными. Поэтому у разных авторов могут сильно различаться смысл отдельных биомов и, естественно, их число (см. также Главу 2).
** Фактически черепаху надо только было перевернуть и развести вокруг неё огонь и еда готова. Не надо было даже вскрывать панцирь, поскольку основная масса съедобного мяса у черепах сосредоточена в лапах. Поскольку размеры черепах достигали нескольких метров в длину, а вес — порядка тонны, при весьма вкусном мясе, добыча эта была приоритетной. В массе гибли черепахи и во время пожаров, частота которых возрастала параллельно с разрушением человеком природных СС.
*** Впрочем, весьма сходны с наконечниками кловис наконечники солютре, известные из Франции и Испании 21—17 тысяч лет назад. Это даже послужило основой для теории заселения Северной Америки из Европы. Однако различие ДНК (Rasmussen et al., 2014) европейцев и индейцев убедительно доказывает несостоятельность этой гипотезы. Сходство наконечников, скорее всего, обусловлено сходным способом охоты и возникло независимо.
**** Максимально зарегистрированные величины достигают 3,4%, как на острове Питкерн (Birdsell, 1957), или даже 4,1% как в конце ХХ века в Кении (Diamond, 2005).
***** Такая плотность в 100 раз меньше средней плотности населения в таких областях, как Ярославская, Ивановская, Нижегородская и в тысячу раз меньше, чем на о. Цейлон, где отряды партизан десятилетиями скрывались в джунглях.
****** Панамский перешеек могла пройти только небольшая группа людей. Поэтому для дальнейшего движения по Южной Америке вновь должен был возникнуть фронт, на что требовалось время.
******* В США в 1970 г было 112 млн. голов крупного рогатого скота. Конечно, не все они находились на вольном выпасе в прериях, хотя корм для тех, кто стоит в коровниках, выращивают, в том числе, и на месте прерий. В любом случае цифры одного порядка, при этом индейцы просто снимали урожай, а американцам приходится ещё и работать. Результатом этой работы является не только продукция мяса, но и стремительная деградация почв.
******** Представление о суровой жизни первобытных людей в непрерывной борьбе с голодом — укоренившийся вымысел. Хотя, бывал и голод. Но животной пищи, в целом, было “полно”. “Вопреки распространенному мнению о том, что древние люди вечно страдали от голода, полевые исследования показали, что даже не в самых изобильных местах первобытные охотники и собиратели обеспечивали себя пищей в относительно короткое время. Конечно, они не жили в вечной праздности, но уже настолько приспособились к окружающей природе, что могли в среднем трудиться не более 2—5 часов в день” (The Origins of Agriculture / Ed. Ch. A. Reed. The Hague Paris, 1977, P. 179. цит по: Гринин, 2003). При этом если добытого хватало на последующие дни, то никто и не спешил продолжать “работу”. Этнограф M.D. Sahlins (1972) одним из первых привлёк внимание к ситуации, при которой первобытные народы могут создавать излишек продукции (пищи и вещей), но не делают и не хотят этого делать, он называл такие социумы обществами изобилия (affluent society). В литературе закрепился термин “парадокс Салинза”. В 50-60-е гг. этнографами было собрано много подобных фактов об.
********* В Африке пройти по следу слона в саванне может даже только что приехавший из Европы турист.