Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Биотическая зональность - Причины зональности биоты - Широтная зональность - Биогеография суши
Распределение различных СС по поверхности Земли является функцией климатических условий, в которых из местной биоты формируются СС. В свою очередь состав местной биот является функцией с одной стороны эволюции, приводящей к возникновению новых групп, а с другой — степени изоляции крупных массивов суши на протяжении кайнозоя и мела и способности разных таксонов к расселению. Различие типов почвы, которые часто также часто называют в качестве причин, влияющих на распределение наземных сообществ, в действительности таковым являться не может. Совершенно очевидно, что почва появляется в результате функционирования СС, в первую очередь растительности, а не наоборот. Более того, как уверяют злые языки, почвоведы часто воспринимают растительность как фактор, мешающий получению почвенных профилей. Почвенные карты часто дают в биогеографических сводках как имеющие некую самостоятельную ценность, а не продукт функционирования экосистем.
“Среди причин макромозаичного распределения степной и лесной растительности... решающее значение принадлежит почве. Леса распространены на... лесных почвах... мощные чернозёмы являются местообитаниями растительности луговой степи” (Вальтер, 1975б: 65 66).
Под пологом леса Quercus ilex “формируется средиземноморская красная почва, или терра-росса, причём вне зависимости от типа материнской породы, которой могут быть известняки или доломиты, мергели, песчаники, базальты, сланцы, гнейсы, граниты или, наконец, аллювиальные отложения” (Вальтер, 1975а: 33).
Климат — “совокупность погодных условий в их среднем годовом и суточном ходе” (Вальтер, 1968: 53). Чисто астрономические причины обуславливают наличие градиента “экватор-полюс”. Однако климат конкретного места определяет и конфигурация морских течений и рельеф местности, а изменение системы морских течений — выраженность этого градиента в геологическом времени. Морские течения и общая схема циркуляции атмосферы рассмотрены в Главе 5. Соседство с водоёмами разного размера, в том числе и внутренними (озёрами, прудами и т. п.) смягчает колебания температуры и увлажнения, их отсутствие — увеличивает. В крайней форме это выражено в океаническом и континентальном типах климатов. Сходное действие оказывают и лесные массивы.
В России используют классификацию советского климатолога Б.П. Алисова (1956), учитывающая особенности циркуляции атмосферы (Рис. Ц1). В ней выделяют четыре основных климатических пояса (экваториальный, тропический, умеренный и полярный), между которыми расположены переходные пояса (субэкваториальный, субтропический и субполярный). Также в классификации используют другое логическое основание: преобладающую циркуляцию воздушных масс. В результате в ней выделяют 16 типов климата (не все возможные сочетания есть в природе): экваториальный пояс (экваториальный климат); субэкваториальный пояс (тропический муссонный климат); тропический пояс (тропический сухой и влажный климаты); субтропический пояс (средиземноморский, субтропический континентальный и субтропический муссонный климаты); умеренный пояс (умеренный морской, умеренно-континентальный, умеренный континентальный, умеренный резко континентальный и умеренный муссонный климаты); субполярный пояс (субарктический и субантарктический климаты); полярный пояс (арктический и антарктический климаты).
За границей используют несколько других классификаций. Широко распространена классификация климатов, предложенная Владимиром Петровичем Кёппеном (Табл. 3.2), тестем автора теории мобилизма Альфреда Вегенера. В её основе лежат режимы температуры и увлажнения. Первоначальный вариант его классификации был опубликован в 1900 г. в работе “Опыт классификации климатов, преимущественно по их отношению к растительному миру”. В позднейших модификациях от растительности остались лишь названия климатов. В последнем авторском варианте, опубликованном в 1936 г., выделено 5 групп климата: тропический, сухой, умеренный, континентальный, полярный, которые были подразделены на 13 климатов и 28 разновидностей. В последней версии классификации (Peel et al., 2007) 29 разновидностей (Рис. Ц2).
Табл. 3.2. Легенда карты климатов по Кёппену (Рис. Ц.2) и определение критериев. По Peel, Finlayson, McMahon (2007).
|
буква шифра |
Описание |
Критерии* |
||
|
1 |
2 |
3 |
||
|
А |
-тропический |
Tcold ≥ 18 |
||
|
f |
-дождевого леса |
Pdry ≥ 60 |
||
|
m |
-муссонный |
Not (Af) & Pdry ≥100-MAP/25 |
||
|
w |
-саванновый |
Not (Af) & Pdry < 100-MAP/25 |
||
|
В |
аридный |
MAP < 10хPthreshold |
||
|
W |
-пустынный |
MAP < 5хPthreshold |
||
|
S |
-степной |
MAP ≥ 5хPthreshold |
||
|
h |
-жаркий |
MAT ≤ 18 |
||
|
k |
-холодный |
MAТ < 18 |
||
|
С |
умеренный |
Thot > 10 & 0 < Tcold < 18 |
||
|
s |
-сухое лето |
Psdry < 40 & Psdry < Pwwet/3 |
||
|
w |
-сухая зима |
Pwdry < Pswet/10 |
||
|
f |
-без сухого сезона |
Not (Cs) or (Cw) |
||
|
a |
-жаркое лето |
Thot ≥ 22 |
||
|
b |
-тёплое лето |
Not (a) & Tmon10 ≥ 4 |
||
|
c |
-холодное лето |
Not (a or b) & 1 ≤ Tmon10 < 4 |
||
|
D |
холодный |
Thot > 10 & Tcold ≤ 0 |
||
|
s |
-сухое лето |
Psdry < 40 & Psdry < Pwwet/3 |
||
|
w |
-сухая зима |
Pwdry < Pswet/10 |
||
|
f |
-без сухого сезона |
Not (Ds) or (Dw) |
||
|
a |
-жаркое лето |
Thot ≥ 22 |
||
|
b |
-тёплое лето |
Not (a) & Tmon10 ≥ 4 |
||
|
c |
-холодное лето |
Not (a, b or d) |
||
|
d |
-очень холодная зима |
Not (a or b) & Tcold < -38 |
||
|
Е |
полярный |
Thot < 10 |
||
|
T |
-тундровый |
Thot > 0 |
||
|
F |
-морозный |
Thot ≤ O |
||
*MAP = среднее годовое количество осадков, MAT — среднегодовая температура, Thot — температура наиболее жаркого месяца, Tcold — температура наиболее холодного месяца, Tmon10 — число месяцев с температурой выше 10°С, Pdry — количество осадков в наиболее сухой месяц, Psdry — количество осадков в наиболее сухой летний месяц, Pwdry — количество осадков в наиболее сухой зимний месяц, Pswet — количество осадков в наиболее влажный летний месяц, Pwwet — количество осадков в наиболее влажный зимний месяц, Pthreshold — переменная, вычисленная по следующему правилу: если 70% MAP приходится на зиму, тогда Pthreshold — 2хMAT, если 70% MAP приходится на лето, тогда Pthreshold = 2хMAT + 28, в противном случае Pthreshold = 2хMAT + 14. Лето (зима) — это наиболее тёплые (холодные) шесть месяцев между октябрём-мартом и апрелем-сентябрём.
Bergeron предложил классифицировать не климаты, а воздушные массы. Для классификации он взял три логических основания, характеризующих воздушные массы: влагосодержание (континентальная, морская), температурные характеристики района происхождения (тропическая, полярная, арктическая или антарктическая, муссонная, экваториальная, верхнеатмосферная) и устойчивость (которую он определял через соотношение температуры воздуха и подстилающей поверхности).
Классификация Thomthwaite основана на балансе воды в почве.
Как видно из этого краткого обзора, типы климата выделяют на основе особенностей циркуляции атмосферы и режимов температуры и увлажнения, а не особенностей биот. Это, впрочем, и не удивительно, поскольку это классификации климатов. Вместе с тем, климат в значительной степени определяет растительность, а, вместе с ней, и всю экосистему. Поэтому биологу необходимо понять, какие характеристики климата являются существенными для биогеографии и существуют ли некие их значения, при которых происходят качественные изменения экосистем. Естественным путём для этого будет проведение биогеографического районирования и последующий анализ связи положения биогеографических границ с характеристиками климата.
Из многочисленных климатических показателей для функционирования наземных сообществ наиболее важны годовые балансы и динамика хода температур и увлажнения. Для наглядности эти данные принято представлять в виде климадиаграмм — графиков, на которых показаны изменения этих параметров по месяцам. В настоящее время наибольшее распространение получила форма климадиаграмм, предложенная известным геоботаником Г. Вальтером в 1960 г. На них показаны ход средних месячных температур в °С и количества осадков в миллиметрах, при этом масштаб 1°=2 мм. Такое соотношение масштабов было подобрано эмпирически, поскольку оно достаточно точно отражает условия увлажнения. В период с достаточным увлажнением кривая температуры расположена ниже кривой осадков, соответствующую часть на диаграмме заполняют вертикальной штриховкой. В засушливый период кривая температуры расположена выше кривой осадков, эту площадь заполняют точечной штриховкой. Во влажных тропиках для осадков, превышающих 100 мм в месяц, масштаб уменьшают в 10 раз, а соответствующую область диаграммы закрашивают чёрным. Это позволяет избежать слишком громоздких диаграмм, уменьшение же точности диаграммы несущественно, поскольку такое количество осадков избыточно и осадки уходят в сток, а не включаются в обмен СС. На климадиаграмме показывают также продолжительность безморозных периодов и абсолютные минимумы и максимумы температур, высоту над уровнем моря и другие параметры. Расшифровка содержания климадиаграммы на конкретном примере дана на Рис. 3.8.
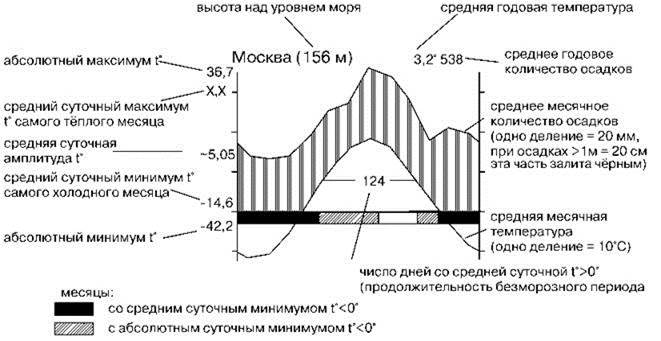
Рис. 3.8. Расшифровка содержания климадиаграммы. По Вальтер (1968), график исправлен по http://pogoda.ru.net/climate/27612.htm.
При всей информативности, существенным недостатком климадиаграмм является то, что они построены на основе данных метеорологических станций, которые размещают на открытых местах, где микроклимат сильно отличается от микроклимата внутри климаксного леса. Больше всего это касается показателей увлажнения (обнажённые участки получают меньше влаги, чем покрытые растительностью). Так, испарение в лесу каменного дуба Quercus ilex в 3—5 раз меньше, чем на открытой местности (Вальтер, 1975: 31). Различны также и амплитуды температуры (внутри леса гораздо меньше), тогда как средние температуры месяцев и года отличаются мало.
Биотическая зональность
Широтная зональность биоты Земли обусловлена астрономическими причинами, приводящими к существованию градиента “экватор—полюс”. Выраженность этого градиента существенно менялась во времени. Поскольку в первую очередь она зависит от системы океанических течений, то факторы, влияющие на градиент “экватор—полюс” рассмотрены при рассмотрении гидрологии окена (Глава 5). Здесь будет рассмотрена только современная зональность.
Разные фитогеографы стремились дать схемы широтной зональности (в зоогеографии суши составлять схемы зональности не принято, зоологи используют схемы ботаников). Вместе с тем, отсутствие общепринятых критериев выделения как типов растительности, так и зон привело к существованию многочисленных схем широтной зональности, порой существенно отличающихся друг от друга. Эти схемы можно объединить в две группы.
“Совершенно необходимо различать естественные и производные, антропогенные, формации, даже если они физиономически сходны. В противном случае изучение их в экологическом аспекте будет невозможно” (Вальтер, 1968: 276).
“Антропогенные “вторичные саванны и злаковники в настоящее время распространены настолько широко, что определяют облик ландшафта... географы часто относят их к коренным” (Вальтер, 1968: 273).
“Вторичные саванны и злаковники в настоящее время распространены настолько широко, что часто определяют облик ландшафта. Поскольку в большинстве случаев трудно доказать их производных характер, географы часто относят их к коренным образованиями. Этим и объясняется выделение зоны коренных “влажных саванн”, где господствуют высокорослые злаки и зоны коренных “сухих саванн” с низкорослыми злаками. Фактически же речь идёт о вторичных группировках, развивающихся на месте определённых категорий лесов” (Вальтер, 1968: 273-274).
“В пустынях Средней Аши сегодня почти невозможно найти следы первоначальной растительности” (Вальтер, 1975: 220).
К первой группе я отнёс схемы идеальных континентов, где показано “идеальное” (от “идея”, а не от “идеал”) распространение растительности на континенте (Рис. 3.9, 3.10). Особо следует подчеркнуть, что большинство авторов, по-видимому, будучи не в силах отойти от географического (ландшафтного) подхода, горные системы выделяет в некие азональные районы, несопоставимые с равнинными районами, или специально оговаривает отсутствие горных экосистем на их схемах, невзирая на отсутствие принципиальных различий биот горных и равнинных районов. Это неадекватное упрощение аналогично биогеографии бентали (Глава 5), где также отдельно и независимо районируют расселение разных глубин).
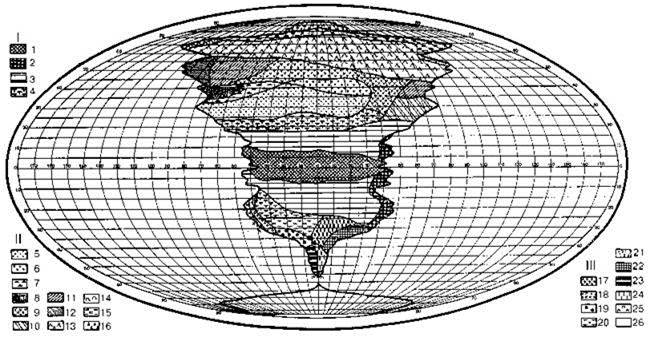
Рис. 3.9. Схема идеального континента, отражающая асимметрию полушарий по Троллю из Г. Вальтера (1968).
I. Тропические зоны: 1 — экваториальный дождевой лес; 2 — тропические дождевые леса, развитые в области пассатных орографических осадков; 3 — тропические (влажные) саванны (см. Рис. 3.2 и сопутствующий текст) и листопадные леса; 4 — тропические колючие степи и колючие леса.
II. Внетропические зоны северного полушария: 5 — сухие пустыни; 6 — холодные внутриматериковые пустыни; 7 — субтропические зимне-зелёные степи; 8 — зона зимних дождей с жарким летом; 9 — зона травяных степей с холодной зимой; 10 — зона влажных муссонных и лавровых лесов с жарким летом; 11 — летнезелёные леса; 12 — океанические летнезелёные листопадные и лавровые леса; 13 — бореальные хвойные леса; 14 — бореальные берёзовые леса; 15 — субарктические тундры; 16 — арктические холодные пустыни.
III. Внетропические зоны южного полушария: 17 — прибрежные пустыни; 18 — пустыни, развитые в пределах пояса “гаруа”; 19 — зона зимних дождей; 20 — субтропические колючие степи (Карру, Монте); 21 — субтропические злаковники; 22 — субтропические дождевые леса; 23 — умеренно холодные дождевые леса; 24 — степи Патагонии и Новой Зеландии; 25 — субантарктические туссоковые луга (антропогенного происхождения см. McWethy et al., 2010) и болота; 26 — антарктическая область материкового льда.
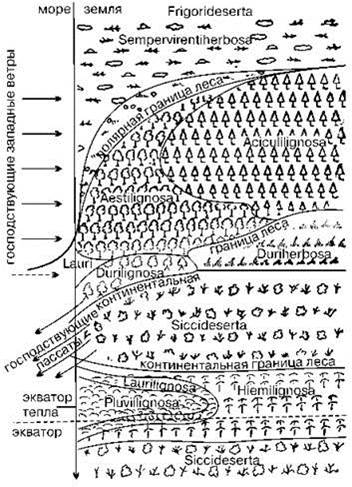
Рис. 3.10. Распределение основных классов и групп формаций на идеальном континенте по В.В. Алёхину и др. (1961).
В схеме использована классификация Брокман-Ероша и Рюбеля.
Различают три основных типа растительности: Lignosa (от лат. lignosus — деревянистый) — древесно-кустарниковый; Herbosa (от лат. herbosus -травянистый) — травянистый; Deserta (от лат. deserta — пустыня) — пустынный,
Их делят на классы формаций. На схеме их 11: Frigorideserta (от лат. frigus — холод) — пустыни, вызванные холодным климатом — холодные пустыни; Sempervirentiherbosa (от лат. semper — всегда, virens — зеленеющий) — вечнозеленые травянистые сообщества; Aciculilignosa (от лат. acuculus — игла, хвоя) — леса и кустарники с листьями в форме игл или чешуи: хвойные леса; Aestilignosa (от лат. aesta — тёплое время года = лето+весна) — летнезелёные (широколиственные) леса; Laurilignosa (от лат. laurus — лавр) — леса и кустарники лаврового типа; Durilignosa (от лат. durus — жесткий) — вечнозеленые жестколистные леса и кустарники; Duriherbosa — с жесткими растениями, например степи; Siccideserta (от лат. siccus — сухой, без вина) — пустыни, вызванные сухостью климата; Hiemilignosa (от лат. hiems — дождливое и холодное время года) — леса и кустарники с листвой в зимнее дождливое время года, листопадные тропические леса; Pluviilignosa (от лат. pluvius — дождливый) — влажные вечнозелёные тропические леса и кустарники.
Ко второй группе я отнёс схемы зонального районирования реальной суши. Схемы на Рис. 3.12 и Рис. Ц.4 на цветной вклейке построены геоботаниками, на Рис. 3.13 и Рис. 3.14 — географами. Для сравнения на Рис. Ц3 на цветной вклейке показана одна из многочисленных схем ландшафтов Земли (также построенная географами). Приведённые схемы весьма различны, причём, за редчайшим исключением авторы схем не обсуждают, чем их не устраивают схемы других авторов, и зачем надо было создавать новую. Причины этого не только в различии методов построения карт, но и в принципиальном различии в теоретических представления о том, что должно быть на этих картах изображено. К любопытным результатам приводит сравнение схем геоботаников и географов. Например схемы геоботаника Г. Вальтера и её модификации географами (Воронов и др., 1985). Эти схемы приведены на Рис. 3.12 и 3.13. Географы считают необходимым выделение саванн в отдельный тип и добавляют биом саванн на карту ботаника Г. Вальтера. В то же время сам Г. Вальтер, специально изучавший этот вопрос, не считает возможным выделять саванны в отдельный биом. Не выделяют саванны и другие геоботаники (Брокман-Ерош, Алёхин). Источник этих разногласий Вальтер (1968) показал на примере того, чем в действительности являются саванны в понимании Тролля (Рис. 3.11).

Рис. 3.11. Ландшафты, которые Тролль (см. Рис. 3.9) относил к “саваннам”. По Вальтеру (1968: 278).
В действительности же это мозаика совершенно различных ассоциаций: чистых злаковников, саванн и больших или меньших лесных массивов. 1 — галерейный саванны; 2 — овражные саванны; 3 — саванны с лесами по береговым валам (“леса бакко”); 4 — галерейно-термитные саванны; 5 — овражно-термитные саванны; 6 — термитные саванны с лесами по береговым валам и “термитными островами”. В термитных саваннах деревья растут только на заброшенных термитниках, где они избегают отрицательных последствий пожаров или затопления.
Термитники чаще всего образуются на месте съеденных деревьев. После полного уничтожения дерева термитники, обычно, отмирают.
Схемы С.М. Разумовского (Рис. Ц4) и Г. Вальтера (Рис. 3.12) построены геоботаниками. При построении схем оба автора пытались восстановить естественную растительность суши (очень сильно антропогенно нарушенную), но каждый делает это на основе своих представлений. Схема Г. Вальтера построена на основе его преимущественно аутэкологических представлений о характере естественной растительности данного места. Несмотря на существенные расхождения в их взглядах на роль ценозов, их схемы более сходны между собой, нежели со схемами географов.
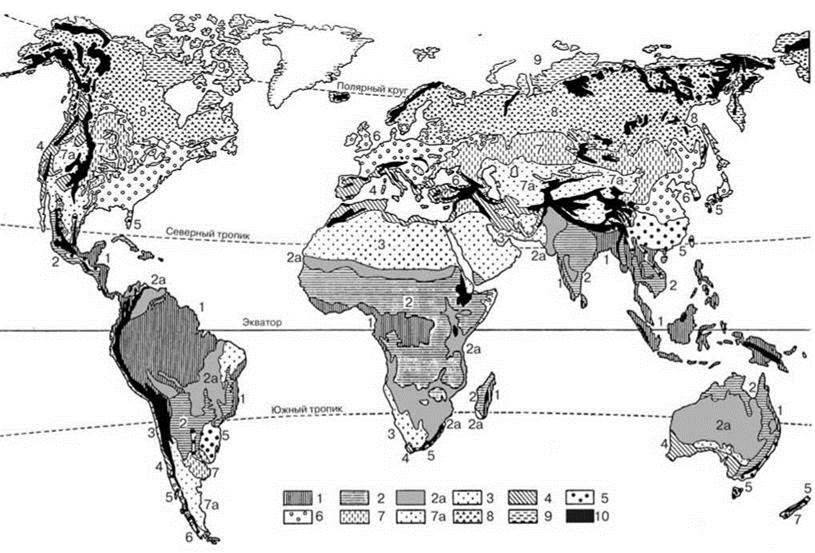
Рис. 3.12. Флористическая зональность Земли по Г. Вальтеру (1975а).
Эдафически обусловленные типы растительности (песчаные пустыни Австралии, обширные заболоченные районы, мангровы и т. д.), а также растительность, измененная деятельностью человека, не учтены.
Тропические и субтропические зоны:
1 — вечнозелёные тропические дождевые леса низменностей и горных склонов (леса пояса туманов); 2 — тропические полувечнозелёные и листопадные леса; 2а — сухие леса, естественные саванны и заросли колючих кустарников, частично злаковники зоны тропиков и субтропиков; 3 — тропические и субтропические полупустыни и пустыни, простирающиеся в сторону полюсов только до 30° (35°) с. и ю. ш. (в других местах — растительность, отмеченная в пункте 7а).
Умеренная и арктическая зоны:
4 — переходная область жестколистной растительности (с зимними дождями), включая аридные области с зимними дождями; 5 — вечнозелёные лиственные леса умеренно теплой зоны или зон с постоянно влажным прохладным климатом; 6 — летнезелёные лиственные леса умеренной зоны с зимним периодом покоя (неморальная зона); 7 — злаковники умеренной зоны (степи, прерии, пампа); 7а — полупустыни и пустыни с холодной зимой (простираются в сторону полюсов от 30-35° с. и ю. ш.), в Тибете — холодная пустыня; 8 — бореальная зона хвойных лесов северного полушария; 9 — арктическая зона тундры Северного Полушария; 10 — горы с различными поясами растительности, включая альпийский и нивальный.
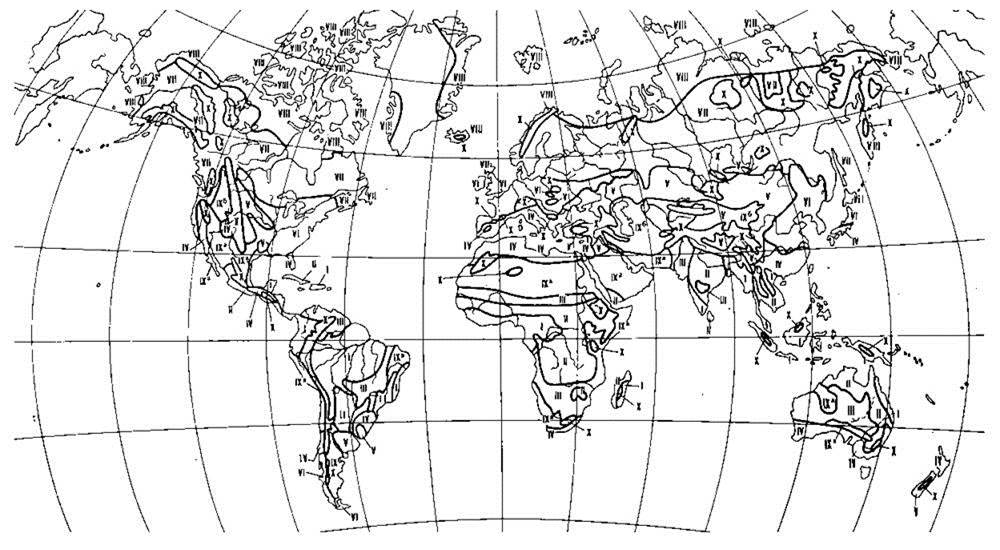
Рис. 3.13. Биомы суши по Г. Вальтеру с изменениями А.Г. Воронова и др. (1985).
Тропические: 1 — влажные экваториальные и тропические леса, 2 — тропические леса, редколесья и колючие кустарники, 3 — саванны. Субтропические: 4 — субтропические леса и кустарники. Умеренные и субполярные: 5 — степи, прерии и их аналоги в южном полушарии, 6 — широколиственные леса, 7 — бореальные хвойные леса, 8 — тундры и их аналоги в южном полушарии, 9 — пустыни (а — тропические и субтропические, б — умеренного пояса), 10 — горные и высокогорные типы.
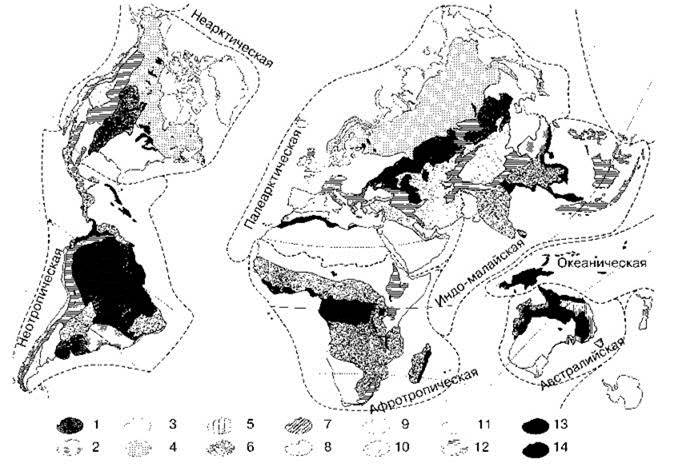
Рис. 3.14. Крупные биомы, соответствующие основным группам экосистем Земли по классификации ЮНЕСКО.
1 — влажные тропические леса, 2 — влажные леса или редколесья субтропического и умеренного поясов, 3 — широколиственные леса или редколесья умеренного пояса и заросли листопадных кустарников субполярных районов, 4 — хвойные леса или редколесья умеренного пояса, 5 — вечнозелёные жестколистные леса, кустарники и редколесья, 6 — сухие тропические или листопадные леса (включая муссонные леса) и редколесья, 7 — тропические степи и саванны, 8 — степи умеренного пояса, 9 — жаркие пустыни и полупустыни, 10 — континентальные пустыни и полупустыни, 11 — тундровые сообщества и арктические пустыни, 12 — смешанные горные и высокогорные системы со сложным зонированием, 13 — смешанные островные системы, 14 — речные и озёрные системы. По План действий..., 1984.
С.М. Разумовский (1999) пошел иным путём. Он начал с установления ботанико-географических провинций, выявил коренные ассоциации их лесных климаксов. Затем он сравнил облик эдификаторов этих климаксных ассоциаций. Оказалось, что их суммарное число для всей Земли составляет лишь несколько десятков видов. Далее он обнаружил, что “эти виды очень легко и естественно распадаются на четыре различных между собой и очень монолитных морфологических типа:
1. Вечнозелёные перистолистные деревья с почками, лишёнными почечных чешуй.
2. Вечнозелёные широколистные деревья с почками, защищёнными почечными чешуями.
3. Летнезелёные широколистные деревья с почечными чешуями.
4. Вечнозелёные деревья с игольчатыми листьями ксероморфной структуры и с почечными чешуями” (Разумовский, 1999: 73).
С.М. Разумовский считал эти зоны естественными биологическими зонами современной растительности Земли: тропическими, субтропическими и широколиственными (неморальными) лесами и тайгой. СС, входящие в одну зону, могут приобрести своё сходство чисто конвергентно и вообще не иметь ничего общего ни в своём составе, ни в происхождении. Сходная ситуация имеется и в таксономии: в воде активно плавающие животные приобретают форму рыбы, но гораздо информативнее классификация не на основе жизненных форм, а филогенетическая, учитывающая то, что одни из этих “рыб” — рыбы, другие — рептилии, третьи — млекопитающие, четвёртые — птицы, пятые — головоногие моллюски и т. д. Из того факта, что в воде они приобрели сходную форму, мало что можно сказать об их внутреннем строении, биохимии, происхождении и т. п. Но все эти данные вполне доступны из филогенетической классификации, построенной на основе их генетического родства. Аналогично, объединение биогеографических регионов по их происхождению гораздо более информативно для понимания их биологических свойств, тогда как зональные классификации лишь показывают многообразие СС, образовавшихся на разной биотической основе, но в сходных абиотических условиях.
Проанализировав данные по связи распределения растительности с температурой и влажностью в 531 точках Земного шара, С.М. Разумовский обнаружил, что для объяснения распределения различных зон растительности достаточно двух параметров: средней температуры самого тёплого месяца и средней температуры самого холодного месяца (Рис. 3.15), увлажнение же оказалось несущественным фактором. Он объяснил это способностью СС эффективно регулировать влажность своих биотопов. Режим увлажнения оказывает влияния не на распределение СС, а лишь на распространённость в данном месте тех или иных сукцессионных стадий и серий. При обсуждении причин возникновения безлесных СС также было показано, что ни режим увлажнения, ни температурные условия сами по себе не являются причинами, обуславливающими возникновение безлесных СС. Поскольку лесные и безлесные ассоциации отличаются прежде всего ролью мегафауны, это подтверждает вывод С.М. Разумовского.
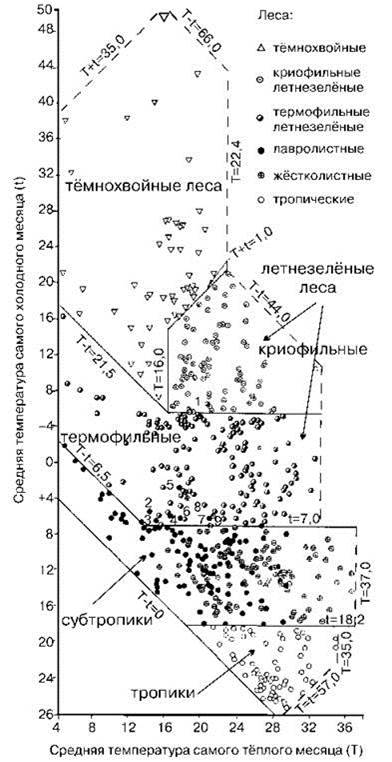
Рис. 3.15. Климатическая ординация растительных зон и подзон. По С.М. Разумовскому (1999).
Г. Вальтер (1985), стоящий на континуалистских позициях, пришёл к тому же выводу об отсутствии безлесного типа растительности как самостоятельной зоны.
Явление листопадности, т. е. синхронизации опадения листьев, характерно для всех зон, за исключением тропической. Г. Вальтер (1968) объясняет его существованием одного неблагоприятного периода. В этих условиях у растений есть два пути адаптации:
1. Листья становятся ксероморфными, приобретая способность переносить неблагоприятный период (вечнозелёные хвойные, жёстколистные виды в областях с сухим летом).
2. Листья опадают в неблагоприятный период.
Растения с ксероморфными листьями могут при наступлении благоприятных условий сразу начать фотосинтез, листопадным же видам необходимо сначала создать листья. Зато затраты на создание листьев у вечнозелёных видов больше. В то же время у вечнозелёных растений листья продолжают функционировать даже при неблагоприятных для фотосинтеза условиях. Это может приводить к таким явлениям, как солнечные ожоги листьев в холодный период (транспирация замедлена, поэтому листья перегреваются).
Листопадные деревья обычны в тропических и субтропических районах с длительным периодом засухи и коротким — дождей (за исключением Австралии) и в умеренных широтах Северного полушария с не очень длительной зимой. Вечнозелёные деревья характерны для тропиков, районов, где осадки выпадают зимой, например в районах со средиземноморским климатом, и для умеренных и высоких широт в районах с длительной зимой. В Австралии листопадные деревья практически отсутствуют, несмотря на существование районов с дефицитом влаги.