БИОЛОГИЯ - справочник для старшеклассников и поступающих в вузы
Обмен веществ и превращение энергии в клетке - УЧЕНИЕ О КЛЕТКЕ - ОБЩАЯ БИОЛОГИЯ
Обмен веществ и превращение энергии — основа жизнедеятельности клетки. Энергетический обмен в клетке и его сущность. Значение АТФ в энергетическом обмене.
Пластический обмен. Фотосинтез. Пути повышения продуктивности сельскохозяйственных растений. Биосинтез белков. Ген и его роль в биосинтезе. Код ДНК. Реакции матричного синтеза. Взаимосвязь процессов пластического и энергетического обмена.
Задание 8
• Повторить имеющийся учебный материал.
• Ответить на вопросы для самоконтроля.
• Выполнить поэтапно контрольные работы № 13-15.
• Проанализировать рисунок 4.
• Проанализировать таблицы 36-41.
• Проанализировать схемы 9-16 и пояснения к ним.
• По словарям основных терминов и понятий проверить знание терминов (Внимание! Эта тема имеет несколько словарей).
Вопросы для самоконтроля (обмен веществ)
• Сформулируйте закон сохранения энергии.
• Назовите главный источник энергии на Земле, определяющий возможность жизни.
• Что такое энергетические уровни в цепи питания?
• Из каких двух противоположных процессов складывается обмен веществ и энергии в организме и в клетке?
• Что называют пластическим обменом в клетке?
• Что такое ассимиляция?
• Расходуется ли АТФ в процессе ассимиляции?
• Какие вещества образуются в процессе ассимиляции?
• Что такие диссимиляция? Почему она проходит поэтапно?
• Что характерно для каждого этапа и как это связано с их названиями?
• Проследите последовательные преобразования крахмала и энергии в процессе диссимиляции (I, II и III этапы).
• Каковы конечные продукты диссимиляции белков, жиров, углеводов?
Обмен веществ и энергии в клетке
Контрольная работа № 13
1. Почему ассимиляция называется пластическим обменом (создаются органические вещества, расщепляются органические вещества)?
2. Почему диссимиляция называется энергетическим обменом (поглощается энергия, выделяется энергия)?
3. Что включает в себя: процесс ассимиляции (синтез органических веществ с поглощением энергии, распад органических веществ с выделением энергии); процесс диссимиляции (синтез органических веществ с поглощением энергии, распад органических веществ с выделением энергии)?
4. Какие процессы, происходящие в клетке, относятся к ассимиляционным (синтез белка, фотосинтез, синтез липидов, синтез АТФ, дыхание)?
5. Чем отличается окисление органических веществ в митохондриях от горения этих же веществ (выделение теплоты, выделение теплоты и синтез АТФ, синтез АТФ; процесс окисления происходит с участием ферментов, без участия ферментов)?
6. Что общего между окислением, происходящим в митохондриях клеток, и горением (образование СO2 и Н2O; выделение теплоты; синтез АТФ)?
7. На каком этапе диссимиляции полимеры расщепляются до мономеров (I, II, III)?
8. Что происходит с глюкозой на II этапе диссимиляции (гликолиз с образованием пировиноградной кислоты; окисление до СO2 и Н2O)?
9. Какой этап диссимиляции называют кислородным (I, II, III) и почему (в процессе реакции к промежуточным продуктам присоединяется кислород; в процессе реакции выделяется кислород)?
10. На каком этапе диссимиляции углеводов синтезируются 2 АТФ (I, II, III); 36 АТФ (I, II, III); АТФ не синтезируется (I, II, III)?
Схема 9. Метаболизм
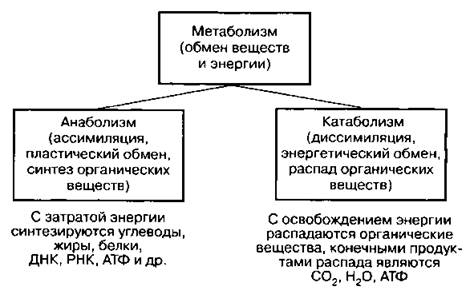
Схема 10. Превращения пировиноградной кислоты в клетках различных организмов
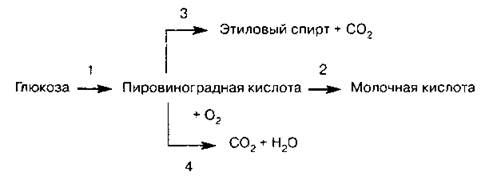
1—2 — анаэробный путь образования молочной кислоты в клетках животных, молочнокислых бактерий и высших растений (в гиалоплазме)
1—3 — спиртовое брожение у дрожжей (анаэробный процесс)
1—4 — дыхание аэробных организмов (в митохондриях)
Пояснения к схеме 11
Превращение веществ и энергии в процессе диссимиляции включает в себя следующие этапы.
I этап — подготовительный: сложные органические вещества под действием пищеварительных ферментов распадаются на простые, при этом выделяется только тепловая энергия.
Белки → аминокислоты
Жиры → глицерин и жирные кислоты
Крахмал → глюкоза
II этап — гликолиз (бескислородный): осуществляется в гиалоплазме, с мембранами не связан; в нем участвуют ферменты; расщеплению подвергается глюкоза:
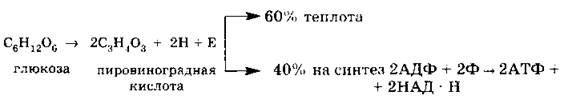
III этап — кислородный: осуществляется в митохондриях, связан с матриксом митохондрий и внутренней мембраной, в нем участвуют ферменты, расщеплению подвергается пировиноградная кислота
![]()
СO2 (диоксид углерода) выделяется из митохондрий в окружающую среду. Атом водорода включается в цепь реакций, конечный результат которых — синтез АТФ. Эти реакции идут в такой последовательности:
Схема 11. Синтез АТФ в митохондрии клетки
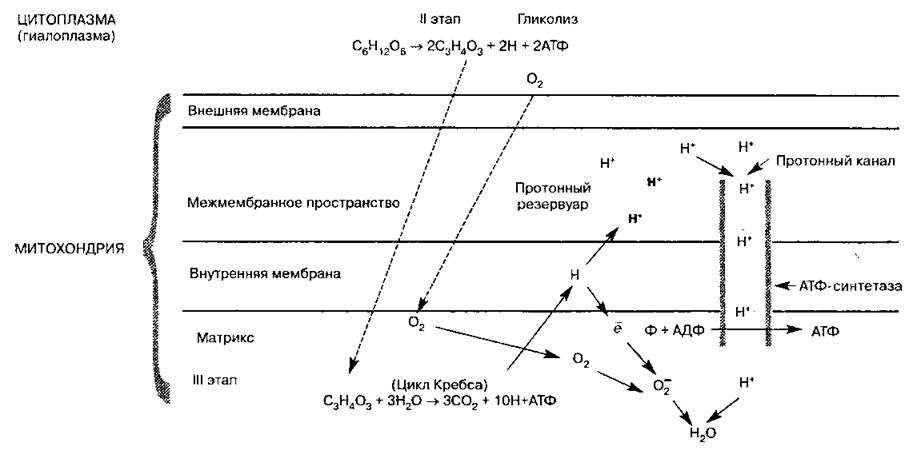
1. Атом водорода Н с помощью ферментов-переносчиков поступает во внутреннюю мембрану митохондрии, образующую кристы, где он окисляется:
![]()
2. Протон Н+ (катион водорода) выносится переносчиками на наружную поверхность мембраны крист. Для протонов эта мембрана, так же как и наружная мембрана митохондрии, непроницаема, поэтому они накапливаются в межмембранном пространстве, образуя протонный резервуар.
3. Электроны водорода е переносятся на внутреннюю поверхность мембраны крист и тут же присоединяются к кислороду с помощью фермента оксидазы, образуя отрицательно заряженный активный кислород (анион):
![]()
4. Катионы и анионы по обе стороны мембраны создают разноименно заряженное электрическое поле, и когда разность потенциалов достигнет 200 мВ, начинает действовать протонный канал. Он возникает в молекулах ферментов АТФ-синтетаз, которые встроены во внутреннюю мембрану, образующую кристы.
5. Через протонный канал протоны Н+ устремляются внутрь митохондрии, создавая высокий уровень энергии, большая часть которой идет на синтез АТФ из АДФ и Ф (АДФ + Ф → АТФ), а сами протоны Н+ взаимодействуют с активным кислородом, образуя воду и молекулярный O2:
![]()
Таким образом, 02, поступающий в митохондрии в процессе дыхания организма, необходим для присоединения протонов Н+. При его отсутствии весь процесс в митохондриях прекращается, так как электронно-транспортная цепь перестает функционировать. Общая реакция III этапа:
![]()
В результате расщепления одной молекулы глюкозы образуются 38 молекул АТФ: на II этапе — 2 АТФ и на III этапе — 36 АТФ. Образовавшиеся молекулы АТФ выходят за пределы митохондрии и участвуют во всех процессах клетки, где необходима энергия. Расщепляясь, АТФ отдает энергию (одна фосфатная связь заключает 40 кДж) и в виде АДФ и Ф (фосфата) возвращается в митохондрии.
Таблица 36. Образование АТФ в ходе диссимиляции
|
Этап диссимиляции, процессы |
Образующиеся макроэнергетические продукты |
Эквивалент АТФ* |
Сколько молекул АТФ образуется |
|
I — Подготовительный |
. |
— |
0 |
|
II — Анаэробный. Гликолиз Образование ацетилкофермента А |
2АТФ |
1 |
2 |
|
2** х 2НАД ∙ Н2 |
3 |
12 |
|
|
III — Аэробный. Цикл Кребса |
2*** х 1АТФ |
1 |
2 |
|
|
2 х 3НАД ∙ Н2 |
3 |
18 |
|
|
2 х 1ФАД ∙ Н2 |
2 |
4 |
|
Итого: |
|
|
38 молекул АТФ образуется при полном окислении 1 молекулы глюкозы |
* Коэффициент, показывающий, сколько молекул АТФ образуется за счет энергии одной молекулы восстановленного переносчика.
** Поскольку из одной молекулы глюкозы образуется две молекулы пировиноградной кислоты, а из каждой молекулы пировиноградной кислоты образуется одна молекула ацетилкофермента А, и при этом восстанавливаются две молекулы НАД.
*** Поскольку за каждый оборот цикла Кребса окисляется одна молекула ацетилкофермента А, то для окисления одной молекулы глюкозы требуется два оборота цикла.
Словарь основных терминов и понятий
Анаболизм (греч. «анаболе» — подъем) — пластический обмен, ассимиляция — одна из сторон обмена веществ. Включает процессы синтеза аминокислот, моносахаридов, жирных кислот, нуклеотидов, а также макромолекул белков, полисахаридов, жиров, нуклеиновых кислот, АТФ. Процесс проходит в три этапа: 1) синтез промежуточных соединений из низкомолекулярных веществ (органических кислот, альдегидов); 2) синтез «строительных блоков» из промежуточных соединений (аминокислот, жирных кислот, моносахаридов); 3) синтез из «строительных блоков» макромолекул белков, нуклеиновых кислот, полисахаридов, жиров. Идет с поглощением энергии и участием ферментов.
Гликолиз (греч. «гликис» — сладкий, «лизис» — растворяю) — бескислородный этап диссимиляции, ферментативный негидролитический анаэробный процесс- распада углеводов до пировиноградной кислоты. Ферменты, ведущие гликолиз, находятся в гиалоплазме (коллоидном веществе цитоплазмы) и не связаны с мембранами. Конечными продуктами гликолиза являются две молекулы пировиноградной кислоты, две молекулы АТФ и две молекулы восстановленного НАД Н2. При невозможности дальнейшего кислородного окисления (у облигатных анаэробов) пировиноградная кислота может окисляться в молочную кислоту (при этом будет затрачиваться одна молекула НАД ∙ Н2 на окисление каждой молекулы пировиноградной кислоты в молочную), этиловый спирт или другие продукты брожения. Если же дальнейшее кислородное окисление возможно, то пировиноградная кислота поступает из цитоплазмы в митохондрии, где претерпевает окислительное декарбоксилирование. Образовавшийся в ходе его ацетил-КоА (ацетилкоэнзим А, ацетилкофермент А) поступает затем в цикл Кребса. Гликолиз — эволюционно наиболее древний путь расщепления глюкозы. У анаэробов он является единственным процессом получения энергии. У аэробов же гликолиз обязательно предшествует кислородному этапу диссимиляции или идет в условиях недостатка кислорода. Гликолиз энергетически значительно менее выгоден, чем кислородное окисление.
Дыхание — цепь физиологических процессов, происходящих в организме растений и животных, при которых поглощается кислород, выделяется двуокись углерода и вода, а также энергия, обеспечивающая жизнедеятельность организма. У животных различают дыхание внешнее (органы дыхания и дыхательные пути) и внутриклеточное (митохондрии), поскольку кислород усваивается только в митохондриях. У растений дыхание осуществляется всеми органами, кислород же усваивается также только в митохондриях клеток. Здесь он включается в электронно-транспортную цепь, присоединяя протоны, кинетическая энергия которых израсходовалась на синтез АТФ. Это кислородный этап диссимиляции (катаболизма), поэтому митохондрии называют дыхательными центрами клеток.
Катаболизм (энергетический обмен, диссимиляция) — одна из сторон метаболизма (обмена веществ), происходящего в любой живой клетке. Катаболизм, или процесс расщепления сложных органических веществ (пищевых, запасных), осуществляется постепенно, в три этапа: I — подготовительный, II — бескислородный (гликолиз), III — кислородный. При этом обязательно участвуют ферменты и выделяется энергия, необходимая для синтеза АТФ и согревания организма (тепловая). Вся энергия, необходимая гетеротрофному организму для жизнедеятельности, получается в результате расщепления органических веществ пищи. Чем больше организм испытывает физических нагрузок, тем больше энергии должна содержать пища и, наоборот, при легкой физической нагрузке пища должна быть малокалорийной.
Метаболизм (греч. «метаболе» — превращение) — обмен веществ — совокупность химических реакций в живой клетке. Единый процесс, свойственный всем живым организмам — животным и растениям. Складывается из двух взаимно противоположных процессов — анаболизма и катаболизма. Значение метаболизма в целом заключается в следующем: 1) поглощение энергии Солнца (растением) и создание органического вещества и пищи (животным); 2) расщепление пищевых веществ до макромолекул — будущих «строительных блоков»; 3) сборка (синтез) из «строительных блоков» белков, жиров, нуклеиновых кислот, углеводов и других веществ; 4) разрушение (расщепление) созданных организмом органических веществ до конечных неорганических — СO2 и Н2O. Метаболизм растений (фотосинтезирующие автотрофы) и животных (гетеротрофы) существенно различается. В живом организме анаболизм и катаболизм должны быть уравновешены. Если один процесс существенно преобладает над другим, то обмен веществ и энергии нарушается и, следовательно, нарушается жизнедеятельность клетки, а затем и всего организма. Метаболизм живой клетки отличается от тепловых процессов неживых тел тем, что живая клетка постоянно черпает энергию из окружающей среды, а та энергия, которая освобождается при расщеплении органических веществ, не рассеивается, а вновь используется на синтез новых веществ, благодаря чему клетка поддерживает свое существование, сопротивляется процессам, ведущим к изменению ее структуры и потере энергии. Неживые же тела теряют свою энергию безвозвратно, так как она переходит в теплоту и рассеивается.
Окислительное фосфорилирование — процесс образования АТФ на клеточных мембранах. Сопряжено с окислением восстановленных переносчиков (например, НАД Н2 до НАД; НАДФ ∙ Н2 до НАДФ) и переносом электронов в электронно-транспортной сети.
Субстратное фосфорилирование — процесс образования АТФ, не связанный с мембранами. В этом случае АТФ образуется за счет того, что фосфатная группа перемещается от фосфорилированного соединения (субстрата) к АДФ (например, образование АТФ при гликолизе).
Хемиосмотическая теория Митчела — учение о механизме преобразования энергии в биологических мембранах. Разработана П. Митчелом в 1961-1966 гг. Согласно этой теории, трансмембранные потенциалы ионов могут служить источником энергии для синтеза АТФ, транспорта веществ и других энергозависимых процессов в клетке. В частности, АТФ синтезируется за счет кинетической энергии протона, проходящего через АТФ-синтетазу (специфический тоннельный белок, пронизывающий мембрану).
Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) — циклическая последовательность ферментативных окислительных превращений девяти три- и дикарбоновых кислот (их декарбоксилирование, дегидрогенизация, изомеризация). Основные этапы этого цикла показаны на схемах 12, 13. Цикл Кребса протекает в матриксе митохондрий в аэробных условиях. Ему предшествует образование ацетил-КоА (ацетилкофермента А, ацетилкоэнзима А) из пировиноградной кислоты (при этом из одной молекулы пировиноградной кислоты образуются также одна молекула углекислого газа и одна молекула восстановленного кофермента НАД ∙ Н2).
![]()
Цикл начинается образованием лимонной кислоты при взаимодействии ацетил-КоА и щавелево-уксусной кислоты и завершается образованием щавелево-уксусной кислоты (для нового цикла), двух молекул углекислого газа, одной молекулы АТФ и четырех молекул восстановленных коферментов (ФАД ∙ Н2 и ЗНАД ∙ Н2). (То есть часть молекул окисляется до конечных продуктов, а часть продолжает «крутиться» в цикле.) Затем (уже вне цикла Кребса) эти восстановленные коферменты окисляются на мембранах митохондрий с образованием АТФ (и воды). Обобщенно превращения, происходящие в цикле Кребса, можно представить следующим образом:

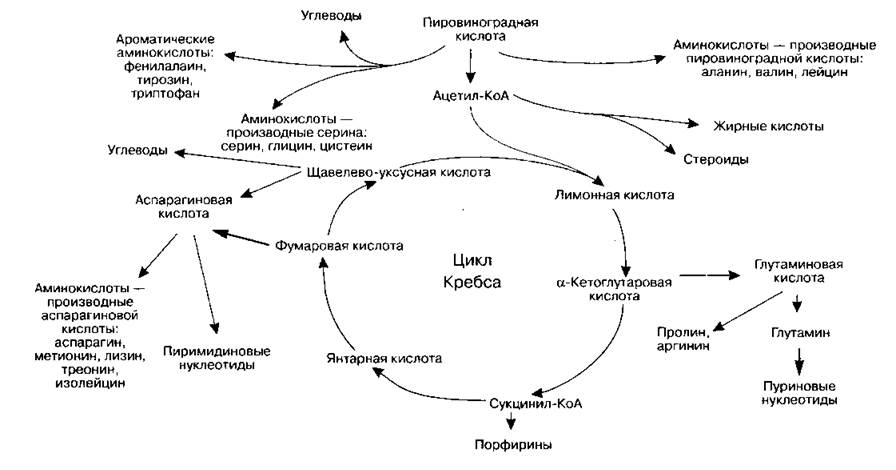
Цикл Кребса широко распространен в клетках животных и растений. Он является основным процессом, обеспечивающим клетку энергией в аэробных условиях. Кроме того, он соединяет воедино различные метаболические пути, поскольку его продукты являются биохимическими предшественниками многих жизненно важных веществ (схема 13).
Схема 12. Цикл Кребса
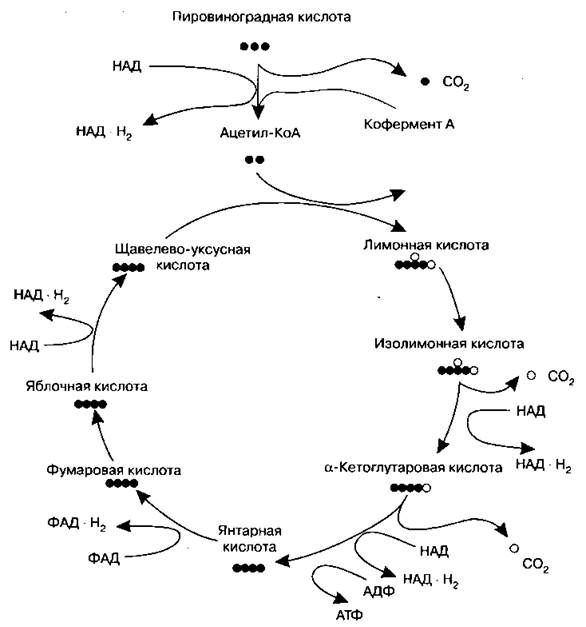
Знак «•» означает один атом углерода. Из данной схемы видно, что для того, чтобы окислиться до углекислого газа (СO2), атом углерода предварительно окисляется до карбоксильной группы (-СООН), обозначенной на схеме знаком «О».
Схема 13. Связь цикла Кребса с различными метаболическими путями
Фотосинтез
Вопросы для самоконтроля
• Почему роль зеленых растений на Земле К. А. Тимирязев назвал космической?
• В какой органелле растительной клетки содержится хлорофилл?
• Когда и у каких растений впервые появился хлорофилл в процессе эволюции?
• Каковы физико-химические свойства хлорофилла?
• Каково внутреннее строение хлоропласта?
• В какой структуре хлоропласта содержится пигмент хлорофилл?
• Как называются мембранные структуры хлоропласта?
• Как называют немембранную часть хлоропласта?
• Чем покрыт хлоропласт и какую форму он может иметь?
• Какие условия необходимы для процесса фотосинтеза?
• Из каких двух фаз (стадий) состоит процесс фотосинтеза?
• Как расходуется энергия возбужденного атома?
• Какие преобразования энергии происходят в хлоропласте?
• Что такое фотолиз воды?
• Какова химическая природа энергетического процесса при фотосинтезе?
• Что образуется в период прохождения световой фазы?
• Какой тип дыхания организмов появился на Земле в связи с возникновением световых реакций фотосинтеза?
• Почему конечные стадии фотосинтеза называют темновыми?
• Какие структуры и какие вещества принимают участие в темновых реакциях фотосинтеза?
• Является ли процесс синтеза углеводов ферментативным?
• В какую фазу фотосинтеза выделяется кислород, уходящий в атмосферу?
• Можно ли считать, что фотосинтез включает в себя два процесса — ассимиляцию и диссимиляцию и почему?
• Какую роль играют углеводы в образовании аминокислот и жирных кислот?
• Можно ли считать растения единственными организмами на Земле, создающими из неорганических веществ органические?
• Какие организмы называют автотрофными?
• Какой способ питания появился на Земле раньше — хемотрофный или фототрофный?
• Откуда получают органические вещества гетеротрофные организмы?
• Чем питаются сапротрофы, паразиты?
• Приведите примеры автотрофных и гетеротрофных организмов.
• Каковы способы питания у зеленой водоросли, белого гриба, гнилостной бактерии, бычьего цепня, волка, человека?

Рис. 4. Строение:
а — хлоропласта, б — лейкопласта, в — хромопласта;
1 — внешняя мембрана, 2 — внутренняя мембрана, 3 — матрикс (строма), 4 — тилакоиды стромы (ламеллы), 5 — грана, 6 — тилакоид граны,
7 — крахмальное зерно, 8 — каротиноиды в каплях липидов, 9 — ДНК, 10 — рибосомы, 11 — разрушающиеся мембранные структуры
Контрольная работа № 14
1. В каких органеллах клетки осуществляется процесс фотосинтеза (митохондрии, рибосомы, хлоропласты, хромопласты)?
2. Где сосредоточен пигмент хлорофилл (оболочка хлоропласта, строма, граны)?
3. Какие лучи спектра поглощает хлорофилл (красные, зеленые, фиолетовые)?
4. При расщеплении какого соединения выделяется свободный кислород при фотосинтезе (СO2, Н2O, АТФ)?
5. В какую стадию фотосинтеза образуется свободный кислород (темновую, световую, постоянно)?
6. Что происходит с АТФ в световую стадию (синтез, расщепление)?
7. На какой стадии в хлоропласте образуется первичный углевод (световая стадия, темновая стадия)?
8. Расщепляется ли молекула СO2 при синтезе углеводов (да, нет)?
9. Какую роль играют ферменты при фотосинтезе (нейтрализуют, катализируют, расщепляют)?
10. Имеется ли хлорофилл у хемосинтезирующих организмов (да, нет)?
11. Какой способ питания у человека (автотрофный, гетеротрофный)?
12. Какие растения создают наибольшую биомассу и выделяют большую часть кислорода (споровые, семенные, водоросли)?
Пояснения к схеме 14. Фотосинтез
Процесс фотосинтеза осуществляется в хлоропластах в два этапа. В гранах (тилакоидах) протекают реакции, вызываемые светом, — световые, а в строме — реакции, не связанные со светом, — темновые, или реакции фиксации углерода.
Световые реакции
1. Свет, попадая на молекулы хлорофилла, которые находятся в мембранах тилакоидов гран, приводит их в возбужденное состояние. В результате этого электроны ![]() сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где и накапливаются, создавая отрицательно заряженное электрическое поле.
сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где и накапливаются, создавая отрицательно заряженное электрическое поле.
2. Место вышедших электронов в молекулах хлорофилла занимают электроны воды ![]() так как вода под действием света подвергается фоторазложению (фотолизу):
так как вода под действием света подвергается фоторазложению (фотолизу):
![]()
Гидроксилы ОН-, став радикалами ОН, объединяются: ![]()
![]() образуя воду и свободный кислород, который выделяется в атмосферу.
образуя воду и свободный кислород, который выделяется в атмосферу.
3. Протоны Н+ не проникают через мембрану тилакоида и накапливаются внутри, образуя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны.
4. При достижении критической разности потенциалов (200 мВ) протоны Н+ устремляются по протонному каналу в ферменте АТФ-синтетаза, встроенному в мембрану тилакоида, наружу. На выходе из протонного канала создается высокий уровень энергии, которая идет на синтез АТФ (АДФ + Ф → АТФ). Образовавшиеся молекулы АТФ переходят в строму, где участвуют в реакциях фиксации углерода.
5. Протоны Н+, вышедшие на поверхность мембраны тилакоида, соединяются с электронами е, образуя атомарный водород Н, который идет на восстановление переносчика НАДФ+: ![]() (переносчик с присоединенным водородом; восстановленный переносчик).
(переносчик с присоединенным водородом; восстановленный переносчик).
Таким образом, активированный световой энергией электрон хлорофилла используется для присоединения водорода к переносчику. НАДФ ∙ Н2 переходит в строму хлоропласта, где участвует в реакциях фиксации углерода.
Реакции фиксации углерода (темновые реакции)
Осуществляются в строме хлоропласта, куда поступают АТФ, НАДФ ∙ Н2 от тилакоидов гран и СО2 из воздуха. Кроме того, там постоянно находятся пятиуглеродные соединения — пентозы С5, которые образуются в цикле Кальвина (цикле фиксации СО2). Упрощенно этот цикл можно представить следующим образом:
1. К пентозе С5 присоединяется СО2, в результате чего появляется нестойкое шестиуглеродное соединение С6, которое расщепляется на две трехуглеродные группы 2С3 — триозы.
2. Каждая из триоз 2С3 принимает по одной фосфатной группе от двух АТФ, что обогащает молекулы энергией.
3. Каждая из триоз 2С3 присоединяет по одному атому водорода от двух НАДФ ∙ Н2.
4. После чего одни триозы объединяются, образуя углеводы
![]()
5. Другие триозы объединяются, образуя пентозы 5С3 → 3С5, и вновь включаются в цикл фиксации СО2.
Суммарная реакция фотосинтеза:
![]()
Схема 14. Фотосинтез

Пояснение к схемам 15, 16. Процесс фотосинтеза
После того как усвоен материал о процессах, происходящих в световую фазу согласно «протонной теории», предлагаем совместить его с теорией «фотосистем». Эти два подхода не только не противоречат друг другу, но, наоборот, дополняют. Преобразование энергии света в энергию химических связей происходит в фотосистемах — ФС (квантососмах), которые являются элементарными структурами, в которых происходит фотосинтез. Они представляют собой пигментно-белковые комплексы, расположенные в мембранах тилакоидов гран. В состав ФС входит от нескольких десятков до сотен молекул хлорофилла, играющих роль антенн, собирающих свет, и лишь несколько из них образуют реакционный центр — РЦ, который и является энергетической «ловушкой» для квантов света. Существует два типа фотосистем — ФСI и ФСII, которые отличаются друг от друга способностью воспринимать свет разной длины волны, что связано с разным сочетанием пигментов, входящих в их состав (см. табл. 37). Легче возбудимы коротковолновые РЦ в ФСII, поэтому именно в них начинается электронно-транспортная цепь (ЭТЦ), которая состоит из переносчиков — цитохромов и передаваемых ими возбужденных электронов. При этом электроны перемещаются в одном направлении, постепенно теряя энергию. Но эта энергия не рассеивается, а идет на синтез АТФ из АДФ и остатка фосфорной кислоты. Этот процесс называется нециклическим фосфорилированием. Электроны, идущие по ЭТЦ, передаются не за пределы мембраны тилакоида, а внутри мембраны к ФСI в ее РЦ. Там энергия принесенного электрона способствует выбиванию более труднодоступного (длинноволнового) электрона ФСI на внешнюю орбиту, так как одной лишь энергии квантов света для этого недостаточно. Выбитый электрон подхватывается специальным переносчиком ферредоксином (из группы цитохромов) и может быть далее отправлен по двум разным путям:
1. Электрон возвратится опять в ЭТЦ и цитохромами будет спущен в ФСI, при этом синтезируется АТФ. Такой путь электрона называется циклическим, а синтез АТФ — циклическим фосфорилированием; он наблюдается в том случае, когда количество свободного НАДФ+ ограничено.
2. Электрон выносится ферредоксином за пределы мембраны тилакоида. На поверхности мембраны создается отрицательный заряд, а электроны присоединяются к переносчику НАДФ+, восстанавливая его, и одновременно с этим подхватывают протоны Н+, вышедшие из протонного канала. На выходе из канала, представленного ферментом АТФ-азой, также происходит синтез АТФ, сопряженный с разностью мембранных потенциалов, но это происходит уже за пределами мембраны.
Схема 15. Процесс фотосинтеза
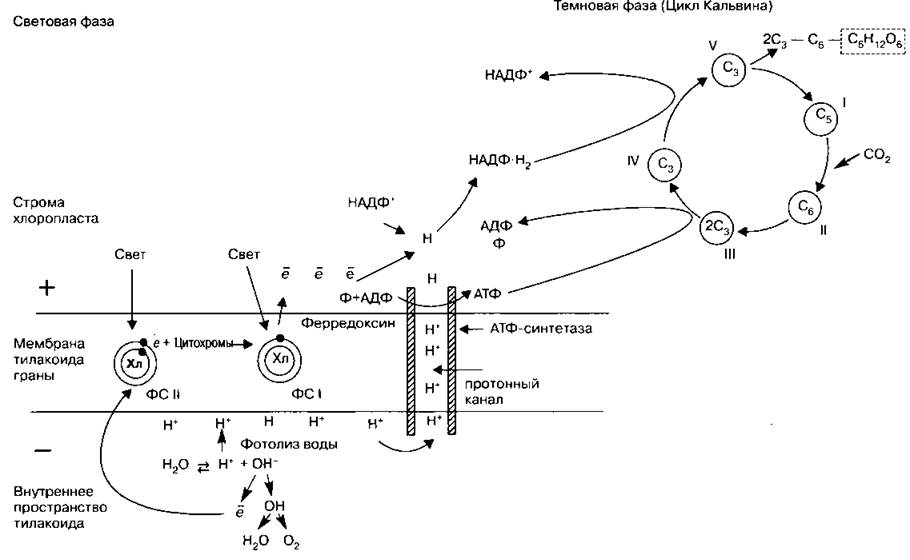
Схема 16. Световая фаза в фотосистемах (по П. Рейвну, Р. Эверту, С. Айкхорн)

Таблица 37. Фотосистемы растительной клетки
|
Признаки |
Фотосистема II (ФСII) |
Фотосистема I (ФСI) |
|
Пигменты |
Хлорофилл а |
Хлорофилл а |
|
Хлорофилл b |
Хлорофилл b |
|
|
Ксантофилл |
Каротин |
|
|
Содержание молекул хлорофилла, шт. |
400 |
250 |
|
Оптимум поглощения света, нм |
680 |
700 |
|
Пигментная система — Р |
P680 |
P700 |
|
Порядок возбуждения электронов |
Первыми возбуждаются более коротковолновые электроны ФСII |
После переноса электрона из ФСII в ФСI возбуждаются более длинноволновые электроны |
|
Возбуждающи й фактор |
Квант света |
Квант света + |
|
Донор электронов |
Вода:
|
ФСII, ФСI |
|
Переносчики электронов |
Цитохромы |
Цитохромы и ферредоксин |
|
Место локализации процесса |
В пределах мембраны тилакоида |
В мембране тилакоида и за ее пределами |
|
Роль |
Перенос электронов на ФСI и возбуждение ее реакционного центра |
1. Возвращение электронов в электронно-транспортную цепь 2. Вынос электронов за пределы мембран тилакоида |
|
Участие в синтезе АТФ |
Синтез АТФ только в ходе нециклического фосфорилирования |
1. Синтез АТФ в ходе и циклического, и нециклического фосфорилирования 2. Восстановление кофермента — переносчика НАДФ+ |
|
Типы фотофосфорилирования (внутри мембраны тилакоида) |
Нециклическое фосфорилирование — однонаправленный поток электронов от ФСII к ФСI; при этом синтезируется АТФ |
1. Циклическое фосфорилирование, при котором поток электронов идет от ФСI в электронно-транспортную цепь и опять возвращается к ФСI; при этом синтезируется АТФ. 2. Нециклическое фосфорилирование, при котором поток электронов идет от ФСII к ФСI и далее за пределы мембраны тилакоида |
Пояснения к таблице 38
В литературе встречаются различные формы записи восстановленной формы НАД (и соответственно НАДФ): НАДвосстан., НАД ∙ Н,НАДН + Н+; НАД ∙ Н2. Как это ни покажется странным, ни одна из них не является ошибочной. Для того чтобы разобраться в этом вопросе, следует рассмотреть механизм восстановления данного кофермента.
Амид никотиновый кислоты, входящий в состав НАД, заряжен положительно, поэтому при взаимодействии НАД с окисляемым органическим веществом, содержащим водород (АН2), субстратом происходит следующая реакция: ![]() Один атом водорода присоединяется к амиду никотиновой кислоты, а от второго атома водорода отрывается лишь электрон, который также присоединяется к молекуле акцептора, делая ее электронейтральной. Акцептированный коферментом водород становится активным, способным легко взаимодействовать с последующими акцепторами. Оставшийся от второго атома водорода протон Н+ временно остается в среде и участвует в химических реакциях позднее. Так образуется восстановленная форма НАД ∙ Н + Н+; сокращенной записью этой формулы является НАД ∙ Н2. Но суть процесса взаимодействия НАД с субстратом заключается не в превращениях НАД, а в активации водорода. Можно даже сказать, что запись НАД ∙ Н2 означает скорее «активированный, реакционноспособный водород», нежели «восстановленный кофермент». Поэтому можно использовать любую из приведенных форм записи восстановленной формы НАД. Однако в том случае, когда надо написать уравнение химической реакции и расставить в нем коэффициенты, удобнее пользоваться формулой НАД ∙ Н2, так как в этом случае не надо «держать в уме» второй протон.
Один атом водорода присоединяется к амиду никотиновой кислоты, а от второго атома водорода отрывается лишь электрон, который также присоединяется к молекуле акцептора, делая ее электронейтральной. Акцептированный коферментом водород становится активным, способным легко взаимодействовать с последующими акцепторами. Оставшийся от второго атома водорода протон Н+ временно остается в среде и участвует в химических реакциях позднее. Так образуется восстановленная форма НАД ∙ Н + Н+; сокращенной записью этой формулы является НАД ∙ Н2. Но суть процесса взаимодействия НАД с субстратом заключается не в превращениях НАД, а в активации водорода. Можно даже сказать, что запись НАД ∙ Н2 означает скорее «активированный, реакционноспособный водород», нежели «восстановленный кофермент». Поэтому можно использовать любую из приведенных форм записи восстановленной формы НАД. Однако в том случае, когда надо написать уравнение химической реакции и расставить в нем коэффициенты, удобнее пользоваться формулой НАД ∙ Н2, так как в этом случае не надо «держать в уме» второй протон.
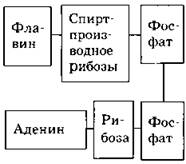
Таблица 38. Переносчики водорода и электронов в клетке
|
Признаки |
Название переносчиков |
||||
|
НАД (никотинамидадениндинуклеотид) |
НАДФ (никотинамидадениндинуклеотидфосфат) |
ФАД (флавинадениндинуклеотид) |
Гемины |
||
|
Классхимических соединений |
Пиридиновые динуклеотиды |
Флавиновые динуклеотиды |
В основном — железопорфирины |
||
|
Состав |
|
|
|
Основу составляет порфириновое ядро, состоящее из четырех пятичленных гетероциклов — пиррольных колец,атомы азота которых соединены в центре с атомом железа |
|
|
В состав каких ферментов входят |
Являются простетическими группами различных дегидрогеназ |
Являются простетическими группами цитохромов (которых насчитывается около 30) |
|||
|
на молекулярном уровне |
Непосредственный акцептор водорода — амид никотиновой кислоты.
Может преобразовываться в НАДФ:
|
Аналогично НАД
Может преобразовываться в НАД:
|
Непосредственный акцептор водорода — флавин
|
Акцептор электрона — окисленное железоFe3+, которое принимает электрон е и становится восстановленным:
(Последний переносчик в дыхательной цепи передает электрон не на атом металла, а на свободный кислород:
|
|
|
В каких процессах клеточного метаболизма участвует |
III (аэробный) этап диссимиляции |
В основном фотосинтез |
III (аэробный) этап диссимиляции |
Электронно-транспортные цепи в ходе ассимиляции и диссимиляции |
|
|
Эквивалент АТФ |
3 АТФ |
3 АТФ |
2 АТФ |
— |
|


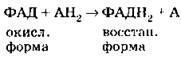
|
Функция в окислительно-восстановительных реакциях |
Перенос водорода. (Активация водорода и передача протонов Н+ и электронов |
Перенос электронов |
||
|
Механизм выполнения своей функции: на клеточном уровне |
Отрывает водород непосредственно от окисляющегося субстрата и передает его другим акцепторам (в дыхательной цепи — ФАД) |
Функционирует аналогично НАД, но в дыхательной цепи не участвует |
Отрывает водород либо от окисляющегося субстрата, либо от предыдущего восстановленного акцептора. (В дыхательной цепи отрывает водород от НАД ∙ Н2 и передает его следующему акцептору.) |
Составляют последовательность переносчиков одних только электронов, которые при диссимиляции получают от предыдущего промежуточного акцептора водорода (в дыхательной цепи), а при фотосинтезе — из фотосистем. (Последний переносчик в дыхательной цепи передает электроны на свободный кислород O2.) |
Схема 17. Темновая фаза фотосинтеза

Каждый прямоугольник □ означает один атом углерода, входящий в углеродный скелет молекулы.
Пояснение к схеме 17. Темновая фаза фотосинтеза
В ходе темновой фазы происходит фиксация углерода, т. е. происходит образование органического вещества (углеводов) из неорганического (углекислого газа). Реакции этой фазы фотосинтеза происходят в строме хлоропластов. В ходе их не используется энергия солнечного света, а используется энергия химических соединений (АТФ и НАДФ ∙ Н2, образовавшихся в световую фазу), поэтому данная фаза и называется темновой.
Фиксация углекислого газа — ключевой момент всей темновой фазы. Акцептором углекислого газа является пятиуглеродное соединение рибулёзо-1,5-дифосфат (РДФ). (Эту реакцию катализирует самый распространенный фермент — рибулёзодифос- фат-карбоксилаза. На его долю приходится примерно 0,2% от массы всего белка, существующего на Земле.) Получающееся шестиуглеродное соединение очень неустойчиво, и оно сразу же распадается на две молекулы трехуглеродного соединения — фосфоглицериновой кислоты (ФГК). Затем ФГК восстанавливается в фосфоглицериновый альдегид (ФГА); в этом процессе участвуют АТФ и НАДФ ∙ Н2. Часть молекул образовавшегося ФГА идет на образование глюкозы (из двух молекул ФГА через серию промежуточных реакций образуется одна молекула глюкозы), а другая часть идет на регенерацию акцептора углекислого газа — рибулёзо-1,5-дифосфата (из пяти молекул ФГА через длинную серию промежуточных реакций образуется три молекулы РДФ). Из данного описания видно, что цепь реакций темновой фазы имеет циклический характер, поэтому она и была названа циклом Кальвина в честь открывшего ее американского ученого М. Кальвина. Цикл Кальвина — наиболее распространенный в природе способ фиксации растениями углекислого газа, хотя имеются и другие, гораздо менее распространенные — путь С4 и кислотный метаболизм толстянковых.
Таблица 39. Сопоставление фотосинтеза и дыхания эукариот
|
Признаки |
Фотосинтез |
Дыхание |
|
Используемыевещества |
Углекислый газ, вода |
Органические вещества, кислород |
|
Итог процесса |
Синтезируется органическое вещество и выделяется свободный кислород (в результате фотолиза воды) |
Разлагается органическое вещество, выделяются углекислый газ и вода |
|
Превращениеэнергии |
Поглощается энергия света, которая преобразуется в энергию химических связей органического вещества (в основном углеводов) |
Высвобождается энергия химических связей органического вещества (в основном углеводов), которая преобразуется в энергию макроэргических связей АТФ |
|
Место образования АТФ |
Хлоропласты |
Митохондрии |
|
Важнейшие этапы процесса |
Световая и темновая фазы (цикл Кальвина) |
Гликолиз и цикл Кребса |
|
Место осуществления процесса |
Хлоропласты растительных клеток |
Гиалоплазма и митохондрии всех эукариотических клеток |
|
Отношение ксолнечному свету |
Происходит только на свету |
Происходит и на свету, и в темноте, так как не нуждается в солнечном свете |
|
Суммарноеуравнение |
|
|
Пояснения к таблице 40
Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических веществ служат процессы окисления различных неорганических веществ: аммиака, сероводорода, серы, водорода и соединений железа (в отличие от фотосинтеза, при котором источником энергии является солнечный свет). Источником водорода для восстановления углекислого газа в ходе хемосинтеза является вода. Хемосинтез был открыт русским ученым С. Н. Виноградским в 1887 г. Важнейшей группой хемосинтезирующих организмов являются нитрифицирующие бактерии, способные окислять аммиак, образующийся при гниении органических остатков, сначала до азотистой, а затем до азотной кислоты:
![]()
Этот процесс сопровождается выделением энергии. Азотная кислота, реагируя с минеральными соединениями почвы, образует нитраты, которые хорошо усваиваются растениями. Бесцветные серобактерии окисляют сероводород и накапливают в своих клетках серу:
![]()
При недостатке сероводорода бактерии производят дальнейшее окисление серы до серной кислоты:
![]()
Железобактерии окисляют двухвалентное железо до трехвалентного:
![]()
Водородные бактерии используют энергию, выделяющуюся при окислении молекулярного водорода:
![]()
Экологическая роль хемосинтеза велика. Нитрифицирующие бактерии широко распространены в природе; они встречаются в почве и в водоемах и участвуют в осуществлении круговорота азота в биосфере. Серобактерии, образуя серную кислоту, способствуют постепенному разрушению и выветриванию горных пород, разрушению каменных и металлических сооружений, выщелачиванию руд и серных месторождений. Серобактерии, окисляющие серу до сульфатов, участвуют в очищении промышленных сточных вод. В результате деятельности железобактерий образуется Fe(OH)3, скопления которого образуют болотную железную руду. Водородные бактерии участвуют в окислении водорода, накапливающегося в результате жизнедеятельности некоторых микроорганизмов, в природных условиях. Они используются также для получения дешевого пищевого и кормового белка, а также для регенерации атмосферы в замкнутых системах жизнеобеспечения (например, система «Оазис-2» была испытана на космическом корабле «Союз-3» в 1973 г.).
Следует хорошо представлять себе различия между хемосинтезом серобактерий и фотосинтезом других групп серобактерий, с одной стороны, и различия между бактериальным фотосинтезом и фотосинтезом зеленых растений — с другой (см. табл. 40).
Вопросы для самоконтроля (синтез белка в клетке)
• Какие условия необходимы для биосинтеза белка?
• Какова роль ДНК в процессе биосинтеза белка?
• Каким образом происходит передача (транскрипция) информации с ДНК на РНК?
• Какова роль иРНК в процессе биосинтеза белка?
• Где образуется и какие функции выполняет тРНК?
• Чему соответствует разнообразие тРНК и как это выражено количественно?
• Что представляет собой антикодон у тРНК?
• Сколько видов аминокислот принимают участие в синтезе белков?
• Каково строение рибосом, где они образуются и размещаются?
• Что такое полисомы?
Таблица 40. Способы питания
|
Показатели |
Автотрофные организмы |
Гетеротрофные организмы |
||
|
фототрофные |
хемотрофные |
сапротрофные |
паразиты |
|
|
Источник получения органического вещества |
Углекислый газ, вода |
Готовые органические вещества мертвых тел |
Готовые органические вещества живых тел |
|
|
Источник получения энергии |
Солнечная радиация, свет |
Реакции окисления неорганических соединений, идущие с выделением энергии |
Реакции окисления белков, жиров, углеводов, идущие с выделением энергии |
|
|
Представители животного мира |
Эвглена зеленая |
Нет |
Большинство животных |
Простейшие, паразитические черви, клещи, насекомые |
|
Представители растительного мира, бактерии, грибы |
Все зеленые растения, окрашенные бактерии (пурпурные и зеленые, содержащие бактериохлорофилл) |
Нитрифицирующие бактерии, серобактерии, железобактерии |
Бактерии брожения, гнилостные бактерии, шляпочные грибы, дрожжевые, плесневые грибы |
Вирусы, фаги, болезнетворные бактерии, паразитические грибы, цветковые растения — паразиты (омела, повилика, заразиха) |
Таблица 41. Особенности основных способов автотрофной ассимиляции
|
Признак |
Группы организмов |
||
|
Хемосинтезирующие серобактерии |
Фотосинтезирующие серобактерии |
Растения |
|
|
Представители |
Бесцветные серобактерии |
Зеленые и пурпурные серобактерии |
Все растения |
|
Тип питания |
Автотрофный |
||
|
Наличие фотосистемы II |
Отсутствует любая фотосистема |
Отсутствует, поэтому кислород не выделяется |
Имеется, поэтому при фотолизе выделяется кислород |
|
Источник энергии для восстановления СO2 |
Окисление H2S (при недостатке H2S — окисление S) |
Солнечный свет |
|
|
Источник водорода для восстановления СO2 |
Вода Н2O |
H2S |
Вода Н2O |
|
Роль H2S |
Источник энергии для восстановления СO2 |
Источник водорода для восстановления СO2 |
Не играет роли |
|
Уравнение реакции (или продукты реакции) |
Различные сложные органические вещества, но не глюкоза, как у фотосинтезирующих организмов |
(Е св — энергия солнечного света) |
(Е св — энергия солнечного света) |

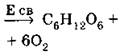
Вопросы для самоконтроля (продолжение)
• Какой процесс происходит в рибосомах и какова роль рРНК?
• Какой процесс при биосинтезе белка называют трансляцией?
• Что представляет собой мономер белковой молекулы?
• Что представляет собой полипептидная цепь?
• Какая структура белка формируется из полипептидной цепи?
• Почему синтез белка в живой клетке называют матричным?
• Чем объясняется многообразие белков и их специфичность?
• Воспроизведите этапы биосинтеза молекулы белка: 1) в хромосоме; 2) на рибосомах; 3) в каналах эндоплазматической сети.
Контрольная работа № 15
1. Какие компоненты клетки непосредственно участвуют в биосинтезе белка (рибосомы, ядрышко, ядерная оболочка, хромосомы)?
2. Какова функция ДНК в синтезе белка (самоудвоение, транскрипция, синтез тРНК и рРНК)?
3. Чему соответствует информация одного гена молекулы ДНК (белок, аминокислота, ген)?
4. Какая структура ядра содержит информацию о синтезе одного белца (молекула ДНК, триплет нуклеотидов, ген)?
5. Какие компоненты составляют тело рибосомы (мембраны, белки, углеводы, РНК, жиры)?
6. Чему соответствует триплет иРНК (аминокислота, белок)?
7. Сколько аминокислот участвуют в биосинтезе белков (100, 30, 20)?
8. Что образуется на рибосоме в процессе биосинтеза белка (белок третичной структуры, белок вторичной структуры, полипептидная цепь)?
9. Где формируются сложные структуры молекулы белка (рибосома, матрикс цитоплазмы, каналы эндоплазматической сети)?
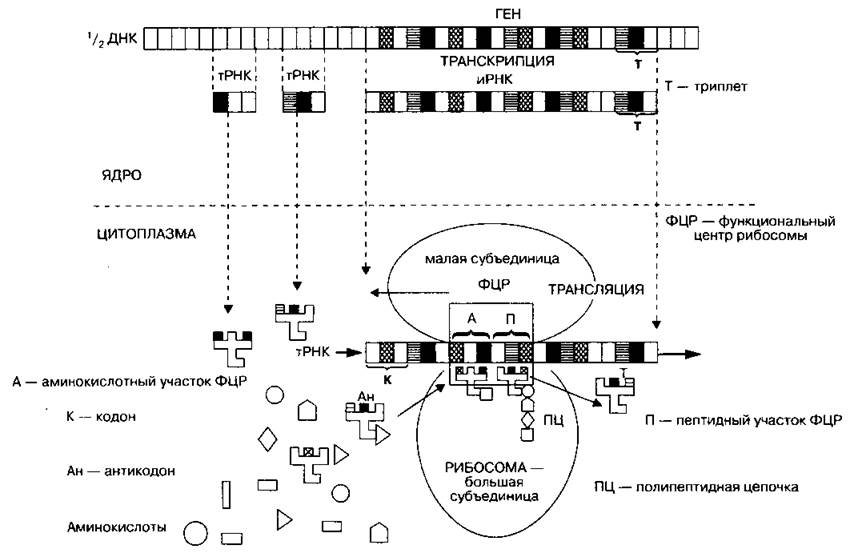
Биосинтез белка
Пояснения к схеме 18
Синтез белка состоит из двух этапов — транскрипции и трансляции.
I. Транскрипция (переписывание) — биосинтез молекул РНК, осуществляется в хромосомах на молекулах ДНК по принципу матричного синтеза. При помощи ферментов на соответствующих участках молекулы ДНК (генах) синтезируются все виды РНК (иРНК, рРНК, тРНК). Синтезируется 20 разновидностей тРНК, так как в биосинтезе белка принимают участие 20 аминокислот. Затем иРНК и тРНК выходят в цитоплазму, рРНК встраивается в субъединицы рибосом, которые также выходят в цитоплазму.
II. Трансляция (передача) — синтез полипептидных цепей белков, осуществляется на рибосомах. Она сопровождается следующими событиями:
1. Образование функционального центра рибосомы — ФЦР, состоящего из иРНК и двух субъединиц рибосом. В ФЦР всегда находятся два триплета (шесть нуклеотидов) иРНК, образующих два активных центра: А (аминокислотный, или аминоацильный) — центр узнавания аминокислоты и П (пептидный, или пептидильный) — центр присоединения аминокислоты к пептидной цепочке.
2. Транспортировка аминокислот, присоединение к тРНК, из цитоплазмы в ФЦР. В активном центре А осуществляется считывание антикодона тРНК с кодоном иРНК, в случае комплементарности возникает связэ, которая служит сигналом для продвижения (скачка) рибосомы вдоль иРНК на один триплет. В результате этого комплекс «кодон иРНК и тРНК с аминокислотой» перемещается в активный центр П, где и происходит присоединение аминокислоты к пептидной цепочке (белковой молекуле). После чего тРНК покидает рибосому.
3. Пептидная цепочка удлиняется до тех пор, пока не закончится трансляция и рибосома не соскочит с иРНК. На одной иРНК может умещаться одновременно несколько рибосом (образуется полисома). Полипептидная цепочка погружается в канал эндоплазматической сети и там приобретает вторичную, третичную или четвертичную структуру. Скорость сборки одной молекулы белка, состоящего из 200-300 аминокислот, составляет 1-2 мин. Формула биосинтеза белка:
ДНК (транскрипция) → РНК (трансляция) → белок
Схема 18. Биосинтез белка
Словарь основных терминов и понятий
Интрон — последовательность нуклеотидов ДНК эукариотов, не несущая генетической информации (гены прокариот не содержат интронов). Одни гены могут содержать несколько десятков интронов, другие могут не содержать их совсем. После транскрипции последовательности нуклеотидов РНК, соответствующие интронам, удаляются в ходе сплайсинга.
Кодон — триплет нуклеотидов. Кодирует один аминокислотный остаток или служит сигналом (для начала или завершения белкового синтеза). Термин употребляется и по отношению к ДНК, и по отношению к РНК.
Оперон — участок ДНК, транскрипция которого осуществляется на одну молекулу информационной РНК под контролем одного специального белка-регулятора. Концепция оперона была предложена в 1961 г. Ф. Жакобом и Ж. Мано для объяснения механизма «включения» и «выключения» генов в зависимости от потребности клетки прокариотического организма в веществах, синтез которых контролируют эти гены. Дальнейшие эксперименты позволили дополнить эту концепцию, а также подтвердили, что оперонная регуляция (т. е. регуляция на уровне транскрипции) является основным механизмом регуляции активности генов у прокариот и ряда вирусов.
В состав оперона прокариот входят структурные гены и регуляторные элементы (не путать с геном-регулятором). Структурные гены кодируют белки, осуществляющие последовательно этапы биосинтеза какого-либо вещества. Этих генов может быть один, два или несколько. Они тесно сцеплены друг с другом и, что самое главное, в ходе транскрипции работают как один единый ген: на них синтезируется одна общая молекула иРНК, которая лишь потом расщепляется на несколько иРНК, соответствующих отдельным генам. Регуляторными элементами являются следующие:
— промотор — участок связывания фермента, осуществляющего транскрипцию ДНК — РНК-полимеразы. Является местом начала транскрипции. Представляет собой короткую последовательность из нескольких десятков нуклеотидов ДНК, с которой специфически связывается РНК-полимераза. Кроме того, промотор определяет, какая из двух цепей ДНК будет служить матрицей для синтеза иРНК;
— оператор — участок связывания регуляторного белка;
— терминатор — участок в конце оперона, сигнализирующий о прекращении транскрипции.
На работу оператора данного оперона влияет самостоятельный ген-регулятор, синтезирующий соответствующий регуляторный белок. Этот ген не обязательно располагается рядом с опероном. Кроме того, один регулятор может регулировать транскрипцию нескольких оперонов. Ген-регулятор также имеет собственный промотор и терминатор. Регуляторные белки бывают двух типов: белок-репрессор или белок-активатор. Они присоединяются к специфическим нуклеотидным последовательностям ДНК оператора, что-либо препятствует транскрипции генов (негативная, отрицательная регуляция), либо способствует ей (позитивная, положительная регуляция); механизмы их работы противоположны. Кроме того, на работу белков-репрессоров могут влиять вещества — эффекторы: соединяясь с репрессором, они влияют на его взаимодействие с оператором.
Схема 19. Оперонная регуляция биосинтеза белков
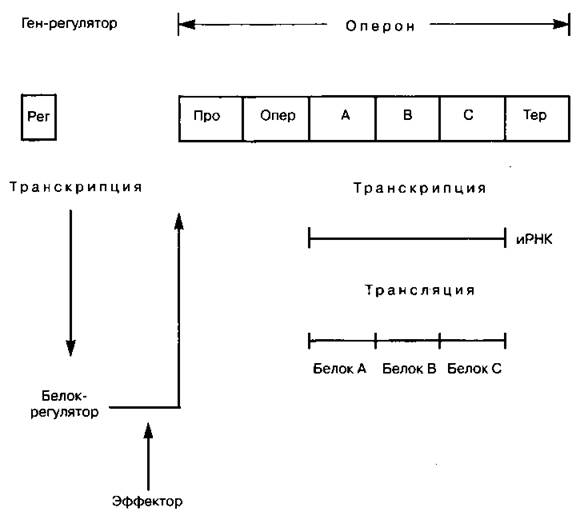
Про — промотор
Опер — оператор
А, В, С, — структурные гены
Тер — терминатор
Таблица 42. Способы регуляции работы оператора
|
Признаки |
Положительная регуляция |
Отрицательная регуляция |
|
Тип белка |
Активатор |
Репрессор |
|
Условие, необходимое для начала транскрипции |
Присоединение белка к оператору |
Отсутствие белка на операторе |
|
Условие, при котором начало транскрипции невозможно |
Отсутствие белка на операторе |
Присоединение белка к оператору |
У эукариот транскрипция осуществляется с участков, подобных оперонам прокариот и также состоящих из регуляторных и структурных генов, однако у оперонов эукариот имеется ряд особенностей.
1. В состав оперона эукариот входит лишь один структурный ген (а не несколько — как у прокариот).
2. Оперон эукариот почти всегда содержит только структурный ген, а прочие гены разбросаны по хромосоме или даже по разным хромосомам.
3. Оперон эукариот состоит из чередующихся друг с другом значащих (экзонов) и незначащих (интронов) участков. При транскрипции считываются как экзоны, так и интроны, а затем в ходе процессинга происходит вырезание интронов (сплайсинг).
У эукариот механизмы регуляции активности отдельных генов и генома в целом довольно сложны, и рассмотрение этих вопросов выходит далеко за рамки школьного курса биологии.
Процессинг (англ. processing — обработка, переработка) — процесс формирования зрелых молекул РНК из их предшественников (пре-РНК). Иными словами, это совокупность реакций, ведущих к превращению первичных продуктов транскрипции (т. е. пре-РНК различных видов) в функционирующие молекулы. Процессинг т- и рРНК в основном сводится к удалению лишних фрагментов с концов молекул. Что касается иРНК, то у эукариот ее процессинг осуществляется многоступенчато. Основными его событиями являются следующие:
— модификация концов молекулы иРНК, в ходе которой к концам молекулы присоединяются специфические короткие последовательности нуклеотидов, обозначающие место начала и место конца трансляции;
— сплайсинг — удаление неинформативных последовательностей РНК, соответствующих интронам ДНК.
У прокариот иРНК не подвергаются процессингу — они способны работать сразу после синтеза.
У всех организмов процессинг РНК происходит в ядре. Для каждого типа молекул он осуществляется специальным ферментом (или группой ферментов).
(Процессингу также могут подвергаться и продукты трансляции, т. е. полипептиды, непосредственно считанные с иРНК. Таким изменениям подвергаются молекулы — предшественники многих белков — пищеварительных ферментов, коллагена, некоторых гормонов, иммуноглобулинов и др., после чего они начинают реально функционировать в организме.)
Сплайсинг — удаление последовательностей нуклеотидов РНК, соответствующих интронам ДНК, и соединение участков с транскрибируемыми последовательностями экзонов. В результате сплайсинга молекулярная масса иРНК уменьшается примерно в 10 раз.
Транскрипция (лат. transcriptio — переписывание) — биосинтез молекул РНК, осуществляемый на молекулах ДНК по принципу матричного синтеза.
Трансляция (лат. translatio — передача) — синтез полипептидных цепей, осуществляемый в рибосомах.
Экзон — информативная последовательность нуклеотидов ДНК. После транскрипции отдельные последовательности нуклеотидов РНК, соответствующие экзонам, в ходе сплайсинга соединяются в единую последовательность, которая и является основной частью зрелой молекулы иРНК. Все экзоны данного гена полностью представлены в молекуле зрелой иРНК, кодирующей первичную структуру белка.