Биология для выпускников школ и поступающих в вузы - Мустафин А. Г. 2015 год
ЭВОЛЮЦИЯ ОРГАНИЧЕСКОГО МИРА - ОБЩАЯ БИОЛОГИЯ
Биологическая эволюция — это историческое развитие живой природы. В ряду сменяющихся поколений от предковых форм до существующих в настоящее время видов возникают наследственные изменения, которые во взаимодействии с факторами среды делают возможным эволюцию в направлении более эффективного развития и размножения организмов в существующих условиях среды. В ходе эволюции осуществляется преобразование видов, возникают новые более приспособленные виды, в то же время происходит исчезновение видов.
Развитие биологии до Дарвина
До 1859 года в естествознании господствовали метафизические взгляды на природу. Все объекты и явления природы рассматривали как неизменные, вне существующих между ними связей. Организмы построены в соответствии с изначальной целесообразностью, т.е. в зависимости от цели, которую поставил Творец. Представление о сотворении живых организмов Богом получило название креационизма. Креационисты (К. Линней, Ж. Кювье) доказывали, что виды живой природы реальны и неизменны со времени своего появления, при этом К. Линней утверждал, что видов существует столько, сколько их было создано во время “сотворения мира”. К концу XVIII в. было описано много животных и растений, проводились попытки их систематизации. Значительный вклад в создание системы природы внес выдающийся шведский естествоиспытатель К. Линней. Он описал более 8 тыс. видов растений и 4 тыс. видов животных. Ученый объединил сходные виды в роды, сходные роды — в отряды, а отряды в классы. Также он ввел принцип двойного наименования (первое родовое и второе видовое название) для обозначения положения определенного вида в системе, например Taenia solium (цепень свиной). В системе Линнея самым крупным таксоном (систематической единицей) был класс, самым мелким — вид. В его классификации было выделено 24 класса растений и 6 — животных. В основу классификации Линней положил принцип иерархичности (соподчиненность) таксонов. Однако его классификация носила искусственный характер, так как основывалась на небольшом числе произвольно взятых признаков и не отражала исторического родства между группами организмов.
Идея исторического развития живой природы и изменяемости организмов зародилась еще у античных мыслителей. К концу XVIII в. в биологии накопился огромный описательный материал, который позволял заключить, что даже внешне далекие виды по внутреннему строению обнаруживают определенные черты сходства, а также что современные виды отличаются от видов, давно живших на Земле. Появившиеся сомнения в неизменности видов привели к возникновению трансформизма - системы взглядов об изменяемости и превращении форм растений и животных под влиянием естественных причин.
Идеи трансформизма нашли дальнейшее развитие в трудах выдающегося французского биолога Ж. Б. Ламарка — создателя первого эволюционного учения. Свои взгляды ни историческое развитие opгaнического мира он изложил в книге “Философия зоологии” (1809). В системе природы Ламарк разместил организмы в восходящем порядке — от простейших до высокоорганизованных существ. Он разделил животных на беспозвоночных и позвоночных, выделил 14 классов, которые распределил на шести ступенях в зависимости от усложнения. Повышение организации от одной ступени к другой было названо им градацией (восхождением). В факте градации ученый увидел отражение хода исторического развития органического мира. По его мнению, эволюция идет на основании внутреннего стремления организмов к прогрессу. Причиной многообразия живого Ламарк считал воздействие различных факторов среды, причем реакции организма на воздействия среды носят целесообразный характер (адекватны изменениям среды) и передаются по наследству.
У организмов, лишенных центральной нервной системы (растения, низшие животные), эти изменения возникают прямым путем. Так, у лютика жестколистного подводные листья сильно рассечены и имеют вид нитей (прямое влияние водной среды), а надводные листья — лопастные (прямое влияние воздушной среды).
У животных, имеющих центральную нервную систему, влияние среды на организм, по Ламарку, осуществляется косвенным путем: условия жизни определяют потребности животного, а значит, действия, привычки и поведение. Вследствие этого одни органы больше и чаще употребляются в работе (упражняются), а другие меньше и реже (не упражняются), причем при упражнении органы развиваются (длинная шея и передние ноги у жирафа, широкие плавательные перепонки между пальцами у водоплавающих птиц и др.), а при неупражнении — недоразвиваются (недоразвитие глаз у крота, крыльев у страуса и др.).
Таким образом, Ж.Б. Ламарк считал, что новые признаки всегда полезны и наследуются. Это представление об изначальной целесообразности любой реакции на измененные условия, так же как и мнение о прямом воздействии окружающей среды на эволюционные процессы и внутреннем стремлении организмов к прогрессу, оказалось ошибочным.
Дарвинизм. Эволюция путем естественного отбора
Фундаментом для создания теории эволюции Ч. Дарвину послужили наблюдения во время кругосветного путешествия на корабле “Бигл”, исследования и обобщение достижений современной ему биологии и селекционной практики. В 1858 году на заседании Линнеевского общества в Лондоне Ч. Дарвин прочитал доклад, содержащий основные положения теории естественного отбора. На том же заседании был прочитан доклад А Уоллеса, высказавшего взгляды, совпадавшие с дарвиновскими. Впоследствии Уоллес признал приоритет Дарвина. В 1859 году вышла книга Ч. Дарвина “Происхождение видов путем естественного отбора, или Сохранение благоприятствуемых пород в борьбе за жизнь”. В ней автор обосновал принцип естественного отбора и представление о борьбе за существование как механизме этого отбора. Основные положении эволюционного учения Ч. Дарвина сводятся к следующим.
Во-первых, для живого характерно наличие изменчивости, причем для эволюции громадное значение имеет наследственная изменчивость, которая носит случайный характер. Вследствие изменчивости признаков и свойств потомки могут отличаться. При благоприятных условиях эти различия могут не играть существенной роли, а при неблагоприятных — каждое мельчайшее различие может стать решающим в том, останется ли этот организм в живых и даст потомство или же не выживет или не оставит потомства.
Во-вторых, каждый вид способен к интенсивному размножению и в каждом поколении возникает гораздо больше особей, чем их может выжить до взрослого состояния на занимаемой территории. Ограниченность жизненных ресурсов препятствует потенциальной возможности интенсивного размножения. Следовательно, значительная часть родившихся гибнет в “борьбе за жизнь”. В процессе жизнедеятельности каждый организм вступает в многообразные отношения с особями внутри вида, других видов и факторами неживой природы. Разнообразные взаимодействия данного организма с объектами живой и неживой природы Дарвин называл борьбой за существование. Он имел в виду “не только жизнь одной особи, но и успех ее в обеспечении себя потомством”. Дарвин выделил три формы борьбы за существование:
1) внутривидовая борьба (взаимоотношения между особями). Ее интенсивность возрастает с увеличением численности и степени специализации вида;
2) межвидовая борьба формируется на базе пищевых отношений между разными видами, а также в конкуренции за места обитания, размножения и т.д. Крайним выражением межвидовых отношений является межвидовая борьба, когда одна форма вытесняет другую или ограничивает ее численность на определенной территории;
3) борьба с неблагоприятными условиями возникает в зависимости от климатических (температура, влажность, освещенность и др.) или почвенных условий, влияющих на жизнедеятельность организма
В-третьих, в результате борьбы за существование происходит гибель или устранение при размножении особей, которые по признакам наименее соответствуют условиям среды обитания. Следствием борьбы за существование является естественный отбор. Естественный отбор, по Дарвину, — это избирательное выживание и размножение наиболее приспособленных организмов. Следует подчеркнуть, что естественный отбор не отбирает более приспособленных, они просто сохраняются в результате элиминации менее приспособленных. В результате этого процесса любая сохранившаяся организация, структура или функция соответствуют состоянию приспособленности друг к другу и к окружающей среде, т.е. оказывается биологически целесообразной. Дарвин впервые показал, что приспособленность на основе отбора не может быть абсолютной, она всегда относительна и адекватна лишь тем условиям среды, в которых виды длительное время существуют.
В-четвертых, Ч. Дарвин считал, что под действием естественного отбора находящиеся в разных условиях группы особей одного вида из поколения в поколение могут накапливать различные приспособления и в итоге образуют новые виды. Он считал, что в основе образования видов лежит принцип дивергенции (расхождения признаков).
Таким образом, результатом отбора будет являться возникновение приспособлений и на этой основе — видового разнообразия. Разнообразные меняющиеся условия среды способствуют эволюции видов в направлении усложнения организации (млекопитающие, насекомые). Если виды обитают долгое время в однородной среде вне жесткой конкуренции, то уровень их организации может оставаться на относительно низком уровне (ланцетники). В постоянно меняющихся условиях среды одни виды, уменьшаясь численно, неизбежно должны погибать и уступать место другим, лучше приспособленным к новым условиям, о чем убедительно свидетельствуют данные палеонтологии.
Синтетическая теория эволюции
В XX веке благодаря данным генетики, экологии, палеонтологии, молекулярной биологии, эмбриологии и ряда других отраслей биологии появились новые возможности для анализа эволюционных преобразований природы. Основанная на данных многих отраслей естествознания (синтетическая) теория выделяет в едином процессе эволюции два взаимосвязанных уровня: микроэволюционный и макроэволюционный.
Микроэволюцией называют начальный этап эволюционных преобразований популяций. Эти преобразования протекают на внутривидовом уровне от возникновения наследственных изменении до формирования адаптаций и возникновения на их основе новых видов.
Макроэволюция изучает процессы эволюции, продолжающиеся после дифференциации новых видов и обусловливающие формирование новых родов, семейств, далее отрядов, классов, типов животных или отделов у растений
Критерии и структура вида. Популяция
Вид — это совокупность особей, характеризующихся общим происхождением, наследственным сходством морфологических, физиологических и биологических особенностей, свободно скрещивающихся и дающих плодовитое потомство, приспособленных к определенным условиям среды, занимающих в природе определенный ареал. В природе виды различаются совокупностью признаков, называемых критериями вида:
• морфологический критерий определяет сходство внешнего и внутреннего строения особей одного вида;
• генетический критерий — одинаковый, типичный для вида, набор хромосом и возможность особей скрещиваться и давать плодовитое потомство;
• физиологический критерий — сходство жизненных процессов;
• биохимический критерий — способность образовывать специфические белки и другие органические вещества;
• экологический критерий — приспособленность к определенным условиям среды,
• географический критерий — распространение в природе в определенном ареале.
Ни один из критериев в отдельности не может служить для определения вида. Охарактеризовать вид можно, только учитывая совокупность всех критериев.
В природе существует генетическая изоляция видов. 1енофонд вида защищен от притока генов особей других видов:
• сроки размножения близких видов могут различаться;
• поведение при спаривании особей разных видов не совпадают;
• если спаривание происходит, то сперматозоиды одного вида не могут проникнуть в яйцеклетку другого;
• межвидовые гибриды обычно нежизнеспособны или бесплодны.
Особи каждого вида внутри ареала распространены неравномерно (рис. 11.1). В его пределах места, благоприятные для их обитания, чередуются с участками, непригодными для их жизни. Поэтому внутри ареала вид распадается на более мелкие единицы - популяции. Особи, населяющие популяцию, связаны между собой сложными взаимоотношениями и общностью эволюционной судьбы. Популяция — это совокупность свободно скрещивающихся особей одного вида в течение большого числа поколений, населяющих определенный ареал и частично изолированных от других популяций. Популяции формируются исторически в определенных экологических условиях. Каждая популяция характеризуется экологически: популяционный ареал, численность особей и ее динамика, возрастная и половая структура. Эволюционируют не отдельные особи, а группы особей, объединенные в популяции. Популяцию считают элементарной эволюционной единицей. Фактор, определяющий единство популяции, и ее относительную обособленность, — свободное скрещивание особей. Внутри популяции образование брачных пар носит случайный характер, частота скрещивания особей внутри популяции гораздо выше, чем между особями соседних популяций. Особи популяции характеризуются генетической общностью и способностью (через скрещивание) свободного обмена генетической информацией при формировании последующих поколений. Совокупность генов всех особей популяции называют генофондом. Важнейшей характеристикой генофонда являются частоты аллелей (генов) и генотипов. Согласно принципу генетического равновесия Харди — Вайнберга (см. главу 9) частоты генотипов по какому-либо гену будут поддерживаться постоянными из поколения в поколение в идеальной популяции. В природных условиях на популяцию действуют внешние и внутренние факторы, влияющие на частоты аллелей и генотипов. Если популяция длительно испытывает значительное давление со стороны каких-либо внешних факторов, то неизбежно произойдет изменение генетического состава популяции. Такое длительное и направленное изменение генотипического состава популяции, ее генофонда получило название элементарного эволюционного явления. Без изменения генофонда популяции эволюционный процесс невозможен. Изменения состава генов популяций происходят под действием множества событий, которые тем или иным путем в состоянии преобразовывать популяции. Тем не менее возможно выделить четыре основных элементарных фактора эволюции: мутационный процесс, популяционные волны, изоляцию, естественный отбор.
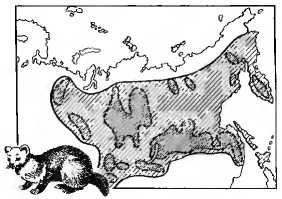
Рис. 11.1. Распространение соболя в Европе:
сплошной линией обозначены границы ареала вида; пунктирной — ареалы популяций
Мутационный процесс
Изменения наследственного материала половых клеток в виде генных, хромосомных и геномных мутаций происходят постоянно в популяции. Общая частота мутаций складывается из частот мутаций отдельных генов. Она колеблется от нескольких процентов у бактерий и одноклеточных эукариот до 10—15% всех гамет одного поколения у многоклеточных организмов. Особое место принадлежит генным мутациям. Они приводят к возникновению серий аллелей и, таким образом, к разнообразию содержания биологической информации. Совокупность аллелей, возникающих в результате мутаций, составляет исходный элементарный эволюционный материал. В процессе видообразования он используется как основа действия других элементарных эволюционных факторов Мутационный процесс постоянно увеличивает генетическую гетерогенность популяций вследствие сохранения рецессивных мутаций в гетерозиготах. Это разнообразие (полиморфизм) усиливается в результате различных генных комбинаций при скрещиваниях. Как показал С. С. Четвериков, популяции насыщены мутациями и обладают широкими возможностями для совершенствования существующих и выработки новых приспособлений при изменении среды. Рецессивные мутации в гетерозиготном состоянии составляют скрытый резерв изменчивости, который может быть использован естественным отбором при изменении условий существования. Но сам мутационный процесс без участия других факторов эволюции не может направлять изменение природной популяции. Он является лишь поставщиком элементарного эволюционного материала, резерва наследственной изменчивости.
Популяционные волны. Дрейф генов
Популяционными волнами называют периодические или апериодические колебания численности организмов в природных популяциях. Это явление распространяется на все виды животных и растений, а также на микроорганизмы. Причинами этих колебаний могут быть различные абиотические и биотические факторы среды. Так, размеры популяций “жертвы” (зайца) растут при снижении давления на них со стороны популяций “хищника” (рыси, лисицы, волка). Отмечаемое в этом случае увеличение кормовых ресурсов способствует росту численности хищников, что в свою очередь интенсифицирует истребление жертвы (рис. 11.2). Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. В условиях роста численности интенсифицируются межпопуляционные миграции особей, что также способствует перераспределению аллелей. Рост количества организмов обычно сопровождается расширением занимаемой территории и выселением некоторых популяций за пределы ареала вида в необычные условия существования. Повышение плотности особей усиливает борьбу за существование.
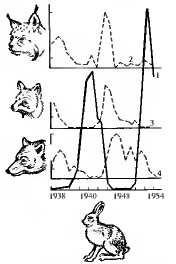
Рис. 11.2. Колебания численности зайцев и хищников в центральных районах европейской части бывшего СССР:
1 — заяц, 2 — рысь; 3 — лисица, 4 — волк
При уменьшении численности может наблюдайся распад крупных популяций. Возникающие малочисленные популяции характеризуются измененными генофондами. В условиях массовой гибели организмов редкие мутантные аллели могут быть генофондом потеряны. Если популяция мала по численности, то в результате случайных событий некоторые особи независимо от своей генетической конституции могут оставить или не оставить потомство, вследствие этого частоты некоторых генов могут резко меняться за одно или несколько поколений. Случайное изменение частот генов в генофонде популяции называют дрейфом генов (рис. 11.3). С дрейфом генов может быть связано явление, называемое эффектом основателя. Когда небольшое число особей в силу разных причин попадают в иные условия существования, то велика вероятность закрепления и распространения в популяции какой- то характерной особенности, имевшейся у одного из основателей популяции. В результате действия эффекта основателя и дрейфа генов популяция может оказаться сильно отличной от той, от которой она ранее отделилась.

Рис. 11.3. Дрейф генов.
Предположим, что одни особи (светлые точки) несут определенный ген, отсутствующий у других особей (темные точки). Если допустить, что в размножении участвует только часть особей популяции, попавших в круг, то по чистой случайности среди них может оказаться только одна особь, несущая данный ген Тогда в следующем поколении этот ген будет встречаться гораздо реже
Изоляция
Ограничение свободы скрещиваний организмов называют изоляцией. Снижая уровень свободною скрещивания, изоляции приводит к увеличению доли близкородственных скрещиваний. Сопутствующая этому гомозиготизация усиливает особенности генофондов популяций, которые создаются вследствие мутаций, комбинативной изменчивости, популяционных волн. Препятствуя снижению межпопуляционных генотипических различий, изоляция является необходимым условием сохранения, закрепления и распространения в популяциях генотипов повышенной жизнеспособности. Различают пространственную и биологическую изоляцию.
Пространственная изоляция связана с территориально-географическими (водные преграды, горные хребты, места, непригодные для жизни, и др.) и экологическими (расселение по разным экологическим нишам) факторами разобщения популяций.
К биологической изоляции могут относиться особенности поведения, изменения строения и физиологической активности сроков размножения и ряда других факторов, препятствующих скрещиванию. После оплодотворения возможны нарушения конъюгации хромосом и ряд других изменений, приводящих к развитию полностью или частично стерильных гибридов, а также гибридов с пониженной жизнеспособностью.
Эволюционное значение разных форм изоляции состоит в том, что она закрепляет и усиливает генетические различия между популяциями.
Изменения частот генов, вызванные изменчивостью, дрейфом генов, популяционными волнами, изоляцией, носят случайный, ненаправленный характер, и даже их совместное действие не приводит к устойчивому осуществлению направленного процесса эволюции. Направляющим фактором эволюции является естественный отбор.
Естественный отбор
Принцип естественного отбора относится к фундаментальным понятиям биологии. Естественный отбор следует понимать, как избирательное выживание и возможность оставления потомства отдельными особями. Биологическое значение особи, давшей потомство, определяется вкладом ее генотипа в генофонд популяции. Отбор действует в популяциях, его объектами являются фенотипы отдельных особей. Фенотип организма формируется на основе реализации информации генотипа в определенных условиях среды. Отбор из поколения в поколение по фенотипам ведет к отбору генотипов, так как потомкам передаются не признаки, а генные комплексы. Таким образом, в современном понимании естественный отбор — это избирательное воспроизведение генотипов, или дифференциальное размножение.
В процессе экспрессии ген может оказывать влияние на многие признаки. В связи с этим в сферу действия отбора могут включаться не только свойства, повышающие вероятность оставления потомства, но и признаки, которые не имеют прямого отношения к воспроизводству. Они отбираются опосредованно в результате корреляций. В ряде случаев отбор может быть направлен на создание взаимных приспособлений видов друг к другу (цветки растений и посещающие их насекомые). Также могут создаваться признаки, вредные для отдельной особи, но обеспечивающие выживание вида в целом (ужалившая пчела гибнет, но, нападая на врага, она сохраняет семью). В целом отбор играет творческую роль в природе, поскольку из ненаправленных наследственных изменений закрепляются те, которые могут привести к образованию новых групп особей, более совершенных в данных условиях существования. Различают стабилизирующий, движущий и разрывающий основные формы естественного отбора.
Стабилизирующий отбор сохраняет то состояние популяции, которое обеспечивает ее максимальную приспособленность в постоянных условиях существования. В каждом поколении удаляются особи, отклоняющиеся от среднего оптимального значения по приспособительным признакам. Он поддерживает средние значения, выбраковывая мутационные отклонения от ранее сформировавшейся нормы. Примером стабилизирующего отбора являются наблюдения за избирательной гибелью домовых воробьев при неблагоприятных погодных условиях. У выживших птиц различные признаки оказывались близкими к средним значениям, а среди погибших эти признаки сильно варьировали. Примером действия отбора в популяциях людей служит большая выживаемость детей со средней массой тела.
Движущий отбор приводит к изменению среднего значения признака в измененных условиях среды. Он обусловливает постоянное преобразование приспособлений видов соответственно изменениям условий существования. Особи популяции имеют некоторые отличия по генотипу и фенотипу. При длительном изменении внешней среды преимущество в жизнедеятельности и размножении может получить часть особей вида с некоторыми отклонениями от средней нормы. Это приведет к изменению генетической структуры, возникновению эволюционно новых приспособлений и перестройке организации вида. Вариационная кривая смешается в направлении приспособления к новым условиям существования (рис. 11.4). Классическим примером этой формы отбора является потемнение окраски бабочки березовой пяденицы в развитых индустриальных районах Англии В сельскохозяйственных районах распространены светлоокрашенные формы; изредка встречающиеся темные формы (мутанты) преимущественно истребляются птицами. Вблизи промышленных центров кора деревьев становится темной из-за исчезновения лишайников, чувствительных к загрязнению атмосферы. Численность темных форм бабочек, менее заметных на стволах деревьев, преобладает.
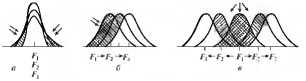
Рис. 11.4. Действие движущей (а), стабилизирующей (б) и разрывающей (в) форм естественного отбора:
F1, F2, F3 — стадии видообразования, стрелками обозначено давление отбора
Разрывающий отбор действует в разнообразных условиях среды, встречающихся на одной территории, и поддерживает несколько фенотипически различных форм за счет особей со средней нормой. Если условия среды настолько изменились, что основная масса вида утрачивает приспособленность, то преимущество приобретают особи с крайними отклонениями от средней нормы. Такие формы быстро размножаются, и на основе одной группы формируется несколько новых (см. рис. 11.4). Основной результат этого отбора заключается в формировании полиморфизма популяции, т.е. наличии нескольких, различающихся по какому-либо признаку групп. Действием разрывающего отбора объясняют образование сезонных рас у некоторых сорных растений. Было показано, что сроки цветения и созревания семян у одного из видов таких растений — погремка лугового растянуты почти на все лето, причем большая часть растений цветет и плодоносит в середине лета. Однако на сенокосных лугах получают преимущества те растения, которые успевают отцвести и дать семена до покоса, и те, которые дают семена в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветушая.
Видообразование
Видообразование — процесс возникновения одного или нескольких новых видов на основе существовавшего ранее — является завершающим этапом микроэволюции. Вид считается качественным этапом эволюционного процесса, гак как он является наименьшей генетически устойчивой надорганизменной системой в живой природе. В ходе видообразования осуществляются в основном следующие процессы:
• возникновение адаптаций к изменению условий среды,
• нарушение связей с родительским видом и постепенное обособление, и на основе изоляции формирование нового вида;
• скрещивание особей нового и старого вида становится невозможным.
Известно два основных пути видообразования: аллопатрическое и симпатрическое.
Новые виды могут появляться в условиях пространственной изоляции популяций, т.е. из популяций, занимающих разные географические ареалы. Такое видообразование называют аллопатрическим (греч. alias — разный, patria — родина), или географическим. В результате длительного разобщения популяций между ними может возникнуть генетическая изоляция, сохраняющаяся даже в том случае, если впоследствии они окажутся вместе. При расселении за пределы ареала исходного вида популяции попадают в иные условия среды обитания, что за счет микроэволюционных процессов может привести к образованию новых видов. Примером дифференциации вида в процессе миграции может быть комплекс популяций и подвидов вида синицы большой. Расселение этого вида из Европы на восток шло двумя путями: северным до Дальнего Востока и южным вокруг Центрально-Азиатского нагорья. На Дальнем Востоке встречаются евроазиатские и восточноазиатские подвиды, которые при совместном обитании не дают гибридов В процессе расселения и преобразования между ними возник репродуктивный барьер. Видообразование путем фрагментации ареала материнского вида хорошо прослеживается на примере возникновения видов ландыша. Лесной ландыш несколько миллионов лет тому назад был широко распространен в Евразии, однако в связи с оледенением его ареал распался на несколько территорий. К настоящему времени сформировалось несколько новых видов.
Симпатрическое видообразование осуществляется в пределах ареала исходного вида. Можно выделить несколько его способов: путем полиплоидии (в роде табака исходное число хромосом равно 12, но имеются формы с 24, 48, 72 хромосомами); путем гибридизации с последующим удвоением хромосом (межвидовые гибриды обычны среди растений, например рябино-кизильник, некоторые виды малины, полыни и др.); путем сезонной изоляции (форель оз. Севан по срокам размножения образует озимую и яровую расы).
Таким образом, в результате влияния эволюционных факторов при ведущей роли естественного отбора идет процесс превращения генетически открытых нестабильных внутривидовых систем популяций в генетически устойчивые видовые системы. Пока между особями разных популяций внутри вида возможны скрещивания и образование плодовитого потомства, осуществляется поток генов между популяциями, вид является единой системой. В результате изоляции прекращается скрещивание, не происходит обмена наследственной информацией и популяции или группы популяций становятся самостоятельными генетическими системами — видами (рис. 11.5).
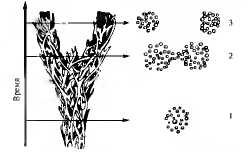
Рис. 11.5. Схема видообразования:
1 — исходным вид, 2 — образование двух подвидов, 3 — два новых вида, каждая отдельная веточка представляет собой популяцию
Макроэволюция
Процесс образования из видов новых родов, из родов — семейств, из семейств — отрядов и так далее называется макроэволюцией. В отличие от микроэволюции, протекающей внутри популяции, макроэволюция — эволюция надвидовая. Между макро- и микроэволюцией нет принципиальной разницы, так как и в том, и в другом случае действуют тe же эволюционные факторы. Тем не менее из-за нарушения характера скрещивания между образовавшимися видами макроэволюция носит необратимый характер. Как указывал Ч. Дарвин, в основе эволюционного процесса лежит дивергенция. Дивергентная эволюция осуществляется на разных уровнях. Она расчленяет вид на популяции. Виды одного рода или роды одного семейства — результат дивергентной эволюции (рис. 11.6). Дивергенция увеличивает разнообразие форм жизни. Морфологические особенности организмов, приобретаемые в процессе дивергенции, имеют некоторую единую основу в виде генофонда родственных форм. Единство происхождения и эволюции подтверждается строением гомологичных органов (рис. 11.7). Гомологичными называют органы, соответствующие друг другу по происхождению и строению независимо от выполняемых функций (конечности позвоночных, видоизменения корня, стебля и листьев у растений).
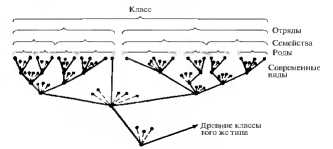
Рис. 11.6. Эволюция путем дивергенции
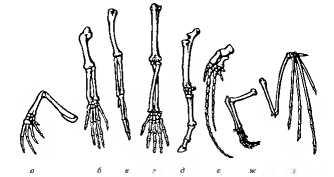
Рис. 11.7. Гомология передних конечностей позвоночных:
а — лягушка, б — ящерица, в — птица, г — обезьяна, д — лошадь, е — кит, ж — кошка, з — летучая мышь
Развиваясь в сходных условиях, неродственные группы могут приобретать сходные признаки. Этот процесс получил название конвергенции (схождения признаков). Конвергентное развитие можно наблюдать у разных групп организмов: внешнее сходство формы тела у акул, ихтиозавров и дельфинов (рис. 11.8), внешнее подобие у многих групп сумчатых и плацентарных млекопитающих и др. Органы, выполняющие одинаковые функции и внешне похожие, но имеющие разное происхождение, называются аналогичными (жабры рака и рыбы, крыло птицы и бабочки, роющие конечности крота и медведки).

Рис. 11.8. Конвергенция у водных животных:
а — акула, б — ихтиозавр, в — дельфин, г — касатка
Отражением эволюционного процесса является систематика организмов. Каждая систематическая группа, стоящая выше по рангу, объединяет группы, стоящие по рангу ниже и имеющие общего предка. Так, род объединяет виды, которые произошли от одного предка и приобрели в результате естественного отбора какие-то различия в фенотипе При построении таксономических систем учитываются признаки родства групп организмов как с ныне живущими группами, так и с уже вымершими. В целом родословное древо отчетливо показывает прогрессивный характер макроэволюции: совершенствование организмов от более простых к более сложным, дивергентный и приспособительный ход эволюции (рис. 11.9).
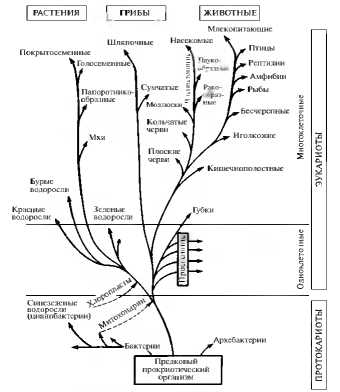
Рис. 11.9. Филогенез основных групп организмов
Направления и пути эволюционного процесса
С момента возникновения жизни развитие живой природы шло от простого к сложному, от низкоорганизованных форм к более высокоорганизованным. В то же время осуществлялись специализация видов, их приспособление к конкретным условиям среды. К этой проблеме обратился А.Н. Северцов. Учение о прогрессе в эволюции было в дальнейшем развито его учеником И. И. Шмальгаузеном. Было предложено из общего понятия эволюционного прогресса выделить биологический прогресс. Биологический прогресс означает успех вида или другой систематической группы в борьбе за существование и характеризуется возрастанием приспособленности особей к окружающей среде. Признаками биологического прогресса являются:
• увеличение численности особей данной систематической группы;
• расширение ареала;
• увеличение количества и разнообразия дочерних групп (популяций и подвидов внутри вида, видов в роде и т.п.)
Все три признака биологического прогресса связаны друг с другом. Увеличение численности приводит к расширению ареала, заселение новых мест обитания приводит к образованию новых популяций, подвидов, видов.
А. Н. Северцов выделил три основных пути достижения биологического прогресса: ароморфозы, идиоадаптации, и дегенерации.
Ароморфоз, или морфофизиологический прогресс, — возникновение в ходе эволюции адаптаций, повышающих уровень организации и эффективность живых существ. Ароморфозы сохраняются в поколениях, приводя к возникновению новых крупных систематических групп — типов, классов. С ароморфозами прежде всего связаны эволюционные преобразования кровеносной, дыхательной, нервной и других систем органов, оказывающих непосредственное влияние на увеличение интенсивности обмена веществ и энергии. Эволюция кровеносной системы от трубчатого сердца у ланцетника к двух-, трех- и четырехкамерному сердцу у позвоночных, сопровождавшаяся обособлением большого и малого кругов кровообращения, шла по пути ароморфозов. Высокий общий уровень организации млекопитающих был достигнут на основе прогрессивного развития кровеносной системы, легких, головного мозга, возникновения живорождения, теплокровности и других ароморфозов. Крупными ароморфозами в развитии растений были переход от размножения спорами к размножению семенами, образование цветка у растений.
Идиоадаптации по А.Н Северцову — частные приспособления видов, позволяющие освоить специфические условия среды. В отличие от ароморфозов идиоадаптации открывают перед организмами возможность биологического прогресса без повышения уровня биологической организации. Например, благодари формированию различных идиоадаптации млекопитающие смогли распространиться не только в различных географических зонах (от тропиков до ледяных пустынь), но и освоить самые разнообразные условия среды (на поверхности суши, в воде, почве, частично в воздухе). Это существенно снизило конкуренцию между видами за пищу, места обитания, причем уровень организации остался тем же. Идиоадаптации к узким, ограниченным условиям среды приводят к специализации группы (бактерии, живущие в горячих источниках; специализация некоторых растений к определенным опылителям и др.). Специализация при быстром изменении условий среды может привести к вымиранию (мезозойские ящеры).
Дегенерация — упрощение строения — часто связана с переходом к пещерному, сидячему или паразитическому образу жизни. Так, пещерные обитатели характеризуются редукцией органов зрения, снижением активности, отсутствием пигментации. Это соответствует дарвиновскому учению, согласно которому эволюция заключается в выживании наиболее приспособленных, а не более высокоорганизованных существ Сидячие организмы (например, асцидии) утратили органы передвижения, хорду, имеющуюся у личиночной стадии. С пассивным образом жизни погонофор связана редукция кишечника, ротового и анального отверстий. Рудиментарные органы — одно из распространенных следствий дегенерации. Упрощение организации обычно сопровождается возникновением различных приспособлений к специфическим условиям жизни. Особенно наглядно это прослеживается на паразитических организмах. У свиного цепня, лентеца широкого и других ленточных червей — паразитов человека и животных нет кишечника, слабо развита нервная система. Однако они отличаются огромной плодовитостью благодаря сильно развитым органам размножения, обладают присосками и крючками, при помощи которых держатся на стенках кишечника своего хозяина. Переход некоторых растений к паразитизму сопровождался снижением активности аппарата фотосинтеза, редукцией листьев до чешуи, преобразованием корней в присоски. Одновременно развивалась сложная система приспособлений к хозяину (химическая сигнализация при поиске растения-хозяина, химический механизм внедрения в ткани хозяина и др.).
Биологический регресс характеризуется снижением численности особей, сужением ареала, постепенным или быстрым уменьшением популяционного и видового многообразия группы. Биологический регресс может привести вид к вымиранию. Исчезли древовидные плауны и хвощи, древние папоротники, большинство древних земноводных и пресмыкающихся. Регрессирующим является род выхухолей, состоящий всего из двух видов, семейство гинкговых и др. Общая причина биологического регресса — отставание в эволюции группы от изменений внешней среды. Быстрое изменение окружающей среды, вызванное деятельностью человека, ведет к увеличению числа видов, переходящих в состояние биологического регресса. Небывалое по скорости наступление человека на природу ведет к столь быстрым изменениям среды, что виды, еще несколько десятков лет назад находившиеся в состоянии биологического прогресса, не поспевая за изменениями, претерпевают биологический регресс. Так, распашка степей ведет к резкому снижению численности целинных видов: сусликов, сурков, степных пищух, журавлей, степных орлов. Минимальные шансы на выживание имеют крупные млекопитающие (особенно хищники), представленные небольшим числом особей, которые живут на небольшой территории, медленно размножаются и не мирятся с соседством человека, например, тигр, гепард, леопард, барс, белый медведь. Напротив, биологического прогресса достигают мелкие плодовитые животные, широко распространенные и легко уживающиеся с человеком. К ним из млекопитающих в первую очередь относится типично городское животное — крыса пасюк, из птиц — ворона, из насекомых — рыжий таракан.
Связь между индивидуальным и историческим развитием организмов
Онтогенез — индивидуальное развитие организма осуществляется в спреде генных условиях внешней среды. В основе индивидуального развития лежит процесс реализации генетической информации, полученной от родителей. Наследственная предопределенность онтогенеза благодаря механизму передачи генетической информации из поколения в поколение закрепляется в процессе исторического развития (филогенеза) данной группы организмов. Любые филогенетические преобразования группы происходят посредством генетически обусловленной перестройки онтогенезов организмов данной группы.
При изучении эмбрионального развития разных групп позвоночных К. Бэр обнаружил, что сначала появляются общие признаки типа, затем последовательно класса, отряда и, наконец, вида (рис. 11.10). Таким образом, эмбрионы разных животных с одним планом строения гораздо более сходны между собой, чем взрослые особи. Исходя из этого Э. Геккель сформулировал биогенетический закон, согласно которому “онтогенез повторяет филогенез”. Другими словами, стадии, которые организм проходит в процессе своего развития, повторяют историческое развитие той группы, к которой он относится. Несмотря на ряд ограничений, эмбрионы или личинки одного типа или класса в общих чертах построены по одному плану. Так, в эмбриогенезе позвоночных появляются или лишь закладываются жаберные щели, сегментированные зачатки мускулатуры на спинной стороне, один круг кровообращения с двухкамерным сердцем. Во взрослом состоянии такое строение сохраняется только у рыб, а по мере развития зародышей других позвоночных появляются признаки амфибий, рептилий, птиц и млекопитающих в соответствии с видовой принадлежностью. Изначальное сходство между зародышами можно расценить как проявление консервативно сохраняющейся наследственной информации, полученной от единых предков.
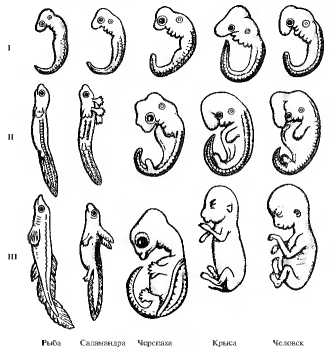
Рис. 11.10. Сходство начальных стадий (I—III) эмбрионального развития позвоночных
В дальнейшем на основании данных сравнительной эмбриологии в работах А.О. Ковалевского, А.Н. Северцова, И.И. Шмальгаузена было показано, что в онтогенезе повторяется строение не взрослых предков, а их зародышей, при этом отдельные стадии могут выпадать. Кроме того, в эмбриональном развитии могут возникать различные морфофизиологические изменения, которые будут определять новые направления филогенеза. Например, у наземных позвоночных зародыш защищен зародышевыми оболочками, которых не было у предковых форм. Следовательно, хотя организмы и сохраняют механизмы развития, унаследованные от предков, но последующие адаптации к иным условиям среды и образу жизни изменяют ход развития.