Биология для выпускников школ и поступающих в вузы - Мустафин А. Г. 2015 год
РАЗВИТИЕ ОРГАНИЧЕСКОГО МИРА - ОБЩАЯ БИОЛОГИЯ
Возраст Земли оценивается примерно 4,6 млрд лет. Первые формы жизни возникли на Земле около 3,2—3,4 млрд лет назад в результате химической эволюции (см. главу 1). На основе анализа находок ископаемых форм в отложениях земных пластов было прослежено историческое развитие живого на Земле. Историю нашей планеты принято делить на эры, эры — на периоды, периоды — на эпохи, эпохи — на века (табл. 12.1).
Таблица 12.1
Геохронологическая шкала. История развития жизни на Земле
|
Эры, их продолжительность и возраст (от начала эры до наших дней), млн лет |
Периоды и их продолжительность |
Животный и растительный мир |
|
Кайнозойская: продолжительность — 67, возраст — 67 |
Антропоген, 1,5 |
Появление, развитие и господство человека. Современный мир животных и растений |
|
Неоген, 23,5 |
Господство млекопитающих, птиц и насекомых. Широкое распространение цветковых растений, особенно травянистых; сокращение флоры голосеменных |
|
|
Палеоген, 42 |
Появление первых приматов (хвостатых лемуров, долгопятов), позднее — парапитеков, дриопитеков. Расцвет насекомых. Продолжается вымирание крупных пресмыкающихся Исчезают многие группы головоногих моллюсков. Господство покрытосеменных растений |
|
|
Мезозойская: продолжительность — 163, возраст — 230 |
Меловой, 70 |
Появление высших млекопитающих и настоящих птиц, хотя и зубастые птицы еще распространены. Преобладают костистые рыбы. Сокращение папоротников и голосеменных. Появление и распространение покрытосеменных |
|
Юрский, 58 |
Господство пресмыкающихся. Появление археоптерикса. Процветание головоногих моллюсков. Господство голосеменных |
|
|
Триасовый, 35 |
Начало расцвета пресмыкающихся Появление первых млекопитающих, настоящих костистых рыб |
|
|
Палеозойская: продолжительность — 340, возраст — 570 |
Пермский, 55 |
Быстрое развитие пресмыкающихся. Возникновение зверозубых пресмыкающихся. Вымирание трилобитов. Исчезновение каменноугольных лесов. Богатая флора голосеменных |
|
Каменноугольный, 75—65 |
Расцвет земноводных Возникновение первых пресмыкающихся. Появление летающих форм насекомых, пауков, скорпионов. Заметное уменьшение трилобитов. Расцвет папоротникообразных. Появление семенных папоротников |
|
|
Девонский, 60 |
Расцвет щитковых. Появление кистеперых рыб Появление стегоцефалов. Распространение на суше высших споровых |
|
|
Силурийский, 30 |
Развитие кораллов Процветание трилобитов. Появление бесчелюстных позвоночных (щитковых) Выход растений на сушу (псилофиты). Широкое распространение водорослей |
|
|
Ордовикский, 60 Кембрийский, 70 |
Процветают морские беспозвоночные. Широкое распространение трилобитов, водорослей |
|
|
Протерозойская: продолжительность — 2000, возраст — 2700 |
Органические остатки редки и малочисленны, но относятся ко всем типам беспозвоночных. Появление первичных хордовых — подтип бесчерепных |
|
|
Архейская: продолжительность — около 1000, возраст 3500 |
Возникновение жизни: прокариоты (бактерии цианобактерии), эукариоты, примитивные многоклеточные |
Доказательства эволюции органического мира
Для обоснования теории эволюции Ч. Дарвин использовал многочисленные доказательства из области палеонтологии, биогеографии, морфологии. Впоследствии были получены факты, воссоздающие историю развития органического мира и служащие новыми доказательствами единства происхождения живых организмов и изменяемости видов в природе.
Палеонтологические находки — едва ли не самые убедительные доказательства протекания эволюционного процесса. К ним относятся окаменелости, отпечатки, ископаемые остатки, ископаемые переходные формы. Среди растений особый интерес представляют псилофиты. Они произошли от водорослей, первыми из растений осуществили переход на сушу. Семенные папоротники — переходная форма между папоротниковидными и голосеменными, а саговниковые — между голосеменными и покрытосеменными. Среди ископаемых позвоночных можно выделить древнейшую группу кистеперых рыб, давшую начало первым земноводным — стегоцефалам. Известны звероящеры, образующие переход между рептилиями и млекопитающими. Связующим звеном между пресмыкающимися и птицами явилась первоптица (археоптерикс). Наличие ископаемых переходных форм доказывает существование филогенетических связей между современными и вымершими организмами и помогает в построении естественной системы и родословного древа растительного и животного мира (см. рис. 11.9).
Установление степени общности и различий в строении организмов является задачей сравнительной анатомии. Внутреннее сходство организмов может показать родство сравниваемых форм. Гомологичные органы (см. главу 11) занимают одинаковое положение в теле животного, развиваются из сходных зачатков и имеют сходный план строения. Типичный пример гомологии — конечности наземных позвоночных животных. Гомологичные органы есть и у растений. Например, усики гороха, колючки кактуса и барбариса — видоизмененные листья
Аналогичные органы свидетельствуют лишь о сходном направлении приспособлений организмов, определяемом в процессе эволюции действием естественного отбора. Жабры рыб, многощетинковых кольчатых червей и водных личинок насекомых (например, стрекоз) аналогичны. Бивни моржа (видоизмененные клыки) и бивни слона (разросшиеся резцы) — типичные аналогичные органы, так как их функции сходны. У растений аналогичны колючки барбариса (видоизмененные листья), колючки белой акации (видоизмененные прилистники) и шиповника (развиваются из клеток коры).
Рудиментарными называются органы, которые закладываются в ходе эмбриогенеза, но в дальнейшем перестают развиваться и остаются у взрослых форм в недоразвитом состоянии. Рудименты — это органы, утратившие свои функции. Например, у муравьедов рудиментарны зубы, у человека — ушные мышцы, кожная мускулатура, третье веко, а у змей — конечности.
Появление у отдельных организмов какого-либо вида признаков, которые существовали у отдаленных предков, но были утрачены в ходе эволюции, называют атавизмами. У человека атавизмами являются хвост, обильный волосяной покров на лице, многососковость и др. Атавизмы и рудименты не несут каких-либо функций, важных для вида, но показывают историческую взаимосвязь между вымершими и ныне существующими родственными формами.
Клеточное строение растений и животных, сходство структурной организации и функционирования клеток, единство принципов хранения, реализации и передачи генетической информации, универсальности генетического кода — наиболее веские доказательства единства органического мира, основанные на данных биологии клетки.
Данные сравнительной эмбриологии имеют большое значение для обоснования эволюционного учения. Сходство гаметогенеза, наличие в развитии одноклеточной стадии — зиготы, сходство зародышей на ранних этапах развития, связь между онтогенезом и филогенезом (см. главу 11) свидетельствуют о единстве мира живых организмов.
Биогеография вносит много доказательств в эволюцию органического мира. Сходные природные зоны Земли заселены различными видами, а разные зоны — сходными. Найти объяснения этим фактам можно только с позиций эволюции. Например, своеобразие флоры и фауны Австралии объясняется обособлением ее в далеком прошлом, в связи с чем развитие животного и растительного мира происходило в изоляции от других материков.
В настоящее время для доказательства эволюционных процессов широко используются методы биохимии и молекулярной биологии, генетики, иммунологии. Так, изучая состав и последовательность нуклеотидов в нуклеиновых кислотах и аминокислот в белках у разных групп организмов и обнаруживая сходство, можно судить об их родстве.
Эволюция клеток
Существует предположение, что все ныне живущие организмы произошли из единственной первобытной клетки. Эта клетка положила начало обмену веществ, процессу клеточного деления и эволюции, которая, в конце концов, создала современную фауну и флору Земли, изменила состав ее атмосферы и сделала ее родиной разумной жизни. На первых этапах эволюции появились прокариоты (см. главу 1). Первичные организмы были анаэробными гетеротрофами и использовали для питания готовые органические вещества, синтезированные абиогенным путем. Далее отбор способствовал возникновению автотрофности (способности к синтезу необходимых органических веществ из неорганических, с использованием в качестве источника энергии либо солнечного света, либо энергии химических связей). Первыми автотрофными фотоеинтетиками были цианобактерии. В результате фотосинтеза в атмосферу начал выделяться кислород, что явилось предпосылкой для возникновения аэробных гетеротрофов, использовавших кислород в процессе обмена веществ.
Около 1 млрд лет назад появились первые эукариотические клетки. Возникновение эукариот связывают с симбиозом прокариотических клеток. Согласно теории эндосимбиоза эукариотическая клетка представляет собой сложную структуру, состоящую из нескольких прокариотических клеток, которые взаимодополняют друг друга в пределах общей клеточной мембраны (см. рис. 1.4). Целый ряд данных свидетельствует о происхождении митохондрий, затем хлоропластов и, возможно, жгутиков от ранних прокариотических клеток, ставших внутренними симбионтами большей по размерам анаэробной клетки. Усложнение строения и функционирования, переход к половому размножению с диплоидной стадией жизненного цикла значительно увеличили эволюционные возможности эукариот, которые смогли достигнуть многоклеточного уровня и сформировать современных животных, растения и грибы.
Эволюция многоклеточных организмов
Преимущества многоклеточности организмов связаны с тем, что все клеточные механизмы и свойства организма оказываются повторенными много раз. Это обеспечивает:
• большую длительность онтогенеза в результате возможности замещения погибших клеток;
• большее число потомков, так как можно образовывать много клеток для размножения;
• значительные размеры и разнообразное строение тела, что обусловливает меньшую зависимость от внешних условий за счет большей стабильности внутренней среды организма.
Царство Животные. Предполагают, что все животные развились от общих предковых форм. Если считать, что наиболее общие стадии онтогенеза повторяют филогенез, то предком многоклеточных следует считать гаструлоподобную форму. Согласно теории гастреи Э. Геккеля, происходило объединение древних жгутиковых в колонию, которая имела шарообразную полость. Затем происходило впячивание одной из сторон колонии с формированием двуслойного гипотетического организма — гастреи (рис. 12.1).
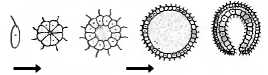
Рис. 12.1. Теория гастреи Геккеля.
Колония жгутиконосцев формирует гипотетический двуслойный организм — гастрею
По мнению И. И. Мечникова, внутренний слой у предковой формы многоклеточных образовался путем миграции специализирующихся на фагоцитозе клеток в полость колонии. Он назвал этот гипотетический организм фагоцителлой. Из гастреи или фагоцителлы можно вывести происхождение губок и кишечнополостных. По плану строения они состоят из двух слоев (эктодермы и энтодермы) и обладают радиальной симметрией. Древние кишечнополостные, видимо, лежат в основе происхождения плоских червей, формирующихся в онтогенезе из трех зародышевых листков и обладающих двусторонней симметрией. Древние ресничные черви дали начало первым вторичнополостным животным — кольчатым червям. Древние морские многощетинковые кольчатые черви, вероятно, являются основой появления типов Членистоногие. Моллюски и Хордовые.
В морях в кембрии палеозоя уже существовали все типы животных. Для большинства из них характерно наличие двусторонней симметрии, третьего зародышевого листка, полости тела, внутреннего (хордовые) или наружного (членистоногие) твердого скелета, прогрессирующая способность к активному передвижению, обособление переднего полюса тела с ротовым отверстием и органами чувств, постепенное совершенствование центральной нервной системы и сенсорного аппарата.
В ордовике появляются рыбообразные бесчелюстные щитковые, отдаленно напоминающие современных круглоротых (миног и миксин), но защищенные мощно развитыми костными пластинами. Рыбы последующих периодов весьма разнообразны: появляются панцирные рыбы (силур), затем хрящевые, двоякодышащие, кистеперые и лучеперые (девон). Наиболее важные ароморфозы — развитие из жаберных дуг подвижных челюстей (обеспечило активный захват добычи), развитие из кожных складок плавников, а затем формирование поясов парных грудных и брюшных конечностей (увеличило маневренность движения в воде) — способствовали перестройке всей организации рыб. Двоякодышащие и кистелерые рыбы посредством пузырей, имеющих связь с пищеводом и снабженных системой кровеносных сосудов, могли дышать атмосферным кислородом. Кистеперые в девоне дали начало первичным земноводным — стегоцефалам.
Выход на сушу первых позвоночных был обеспечен преобразованием плавников в конечности наземного типа, воздушных пузырей в легкие. Освоение суши пресмыкающимися обеспечило в карбоне: сухие ороговевшие покровы, внутреннее оплодотворение, защитные оболочки зародыша и богатая желтком яйцеклетка. От примитивных рептилий через зверозубых ящеров (триас) появились первые млекопитающие. Позднее также от одной из ветвей пресмыкающихся появляются (юра) зубатые птицы (археоптерикс), а затем современные птицы (мел). Теплокровность, четырехкамерное сердце, одна дуга аорты (создает полное разделение большого и малого кругов кровообращения), интенсивный обмен веществ обеспечивают расцвет млекопитающих и птиц. В конце мезозоя появляются плацентарные млекопитающие, позднее (палеоген) от насекомоядных обособляется отряд Приматы. Важнейшим событием антропогена было появление человека.
Параллельно эволюции позвоночных шло развитие беспозвоночных. Переход из водной в воздушную среду осуществился у паукообразных и насекомых на основе совершенствования твердого наружного скелета, членистых конечностей, органов выделения, нервной системы и поведенческих реакций, формирования органов воздушного дыхания. Среди моллюсков выход на сушу наблюдался значительно реже и не приводил к такому разнообразию видов, какое наблюдалось у насекомых. В целом наиболее высокоразвитые беспозвоночные (насекомые и головоногие моллюски) уступают по сложности высшим классам хордовых животных.
Царство Растения. У истоков низших растений находятся примитивные жгутиковые. Видимо, переход к многоклеточности неоднократно осуществлялся через колониальные формы одноклеточных и нитчатых форм зеленых водорослей к многоклеточным зеленым, бурым и красным водорослям. В это же время, вероятно, появились первые водоросли, прикрепленные к дну. У некоторых бурых водорослей в цикле развития начинает преобладать спорофит с его эволюционными преимуществами, обусловленными диплоидностъю; совершенствуется строение таллома (расчленение, многослойность).
В конце силура происходили крупные горообразовательные процессы, повлекшие гибель множества животных и растений, оказавшихся на суше. В прибрежных областях, в условиях периодического заливания водой из многоклеточных водорослей развились первые обитатели суши — псилофиты. Слоевище псилофитов имело ткани (покровные, механические, проводящие), сложную структуру осевого органа — таллома. Псилофиты дали начало наземным высшим растениям: споровым (мохообразным, плаунам, хвощам, папоротникам) и через семенные папоротники семенным растениям (голо- и покрытосеменным). Дальнейшая эволюция растений в наземных условиях шла по пути дифференциации вегетативных органов (появления корней, листьев, более сложного ветвления стебля), развития покровных тканей с толстостенными, содержащими восковидные вещества клетками, совершенствования проводящей системы (переход от трахеид к сосудам). Гаметофит перемещается на спорофитное материнское растение; с развитием пыльцы перед оплодотворением появляется опыление.
Царство Грибы. Обособленная группа эукариотических гетеротрофных организмов с сапрофитным, паразитическим или симбиотическим образом жизни. Видимо, ведут происхождение от гетеротрофных примитивных эукариот, близких к предковым формам 1лавной линии царства животных.