БИОЛОГИЯ пособие для поступающих в вузы
Том ІІ ботаника, анатомия и физиология, эволюция и экология 2018 год
Органический мир состоит из животных и растений. Ископаемые и современные животные подразделяются на две группы: беспозвоночных и позвоночных. Беспозвоночные в свою очередь делятся еще на две группы организмов: одноклеточных (составляют один тип простейших) и многоклеточных (все остальные типы).
Первые одноклеточные существа, появившиеся в глубокой древности, в дальнейшем развивались двумя различными путями: из одних произошли растения, из других — животные. Основным отличием растений от животных является принцип питания организмов. Клетки растений способны сами создавать органические вещества на свету из веществ неорганических — углекислого газа и воды. Строятся эти вещества с помощью хлоропластов, наполненных хлорофиллом и плавающих в цитоплазме клеток растений. Такой тип питания, характерный для растений, называется автотрофным. Животные таким способом не могут вырабатывать органические вещества. В их клетках отсутствуют хлоропласта, и поэтому они потребляют уже готовые органические вещества, созданные растениями. Такой тип питания называется гетеротрофным и свойствен животным организмам и грибам. Таким образом, животные питаются либо растениями, либо другими травоядными животными. Некоторые простейшие одноклеточные организмы обладают и автотрофным и гетеротрофным типом питания. Особенно интересны одноклеточные, питающиеся одновременно двумя способами. Таков жгутиконосец — эвглена зеленая. Это позволяет считать, что такие простейшие стоят недалеко от общих истоков происхождения как растительного, так и животного мира. Те потомки простейших одноклеточных, у которых сохранился хлорофилл, по не было приспособлений для поглощения чужих белков, дали начало миру растений. Другие потомки приобрели способность поглощать и переваривать чужие белки, углеводы и жиры, но в их клетках хлорофилловые зерна не развивались. Такие организмы дали начало миру животных.
Под макроэволюцией подразумевают события, происходящие в геологическом масштабе времени и состоящие в развитии признаков, по которым различаются такие группы, как род, семейство, отряд, класс и тип. Макроэволюцию нельзя изучать путем прямых наблюдений или экспериментов. Вскрыть эти закономерности помогают данные палеонтологии, сравнительной эмбриологии, а также молекулярной биологии и генетики.
Микроэволюция изучает изменения, происходящие внутри вида и приводящие к его дифференцировке, завершаясь видообразованием.
Происхождение многоклеточных организмов
В настоящее время не вызывает сомнения, что многоклеточные организмы произошли от одноклеточных. Из существующих теорий происхождения многоклеточных основными являются теория гастреи Э. Геккеля и теория паренхимеллы И. И. Мечникова. Обе теории основываются на биогенетическом законе Геккеля—Мюллера: “Онтогенез является кратким и быстрым повторением филогенеза, обусловленным наследственностью и приспособлением” (рис. 6.10).
Рис. 6.10. Происхождение многоклеточных животных: верхний ряд — фазы эмбрионального развития многоклеточных животных: 1 — образование двух бластомеров; 2 — полное равномерное дробление; 3 — бластула; 4 — паренхимула; 5 — гаструла; нижний ряд — животные, организация которых соответствует определенной фазе индивидуального развития: 1 — делящаяся эвглена; 2 — колония эудорины; 3 — колония вольвокса; 4 — фагоцителла (гипотетический предок многоклеточных животных); 5 — гидра
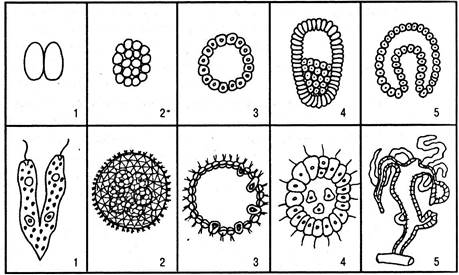
Теория гастреи. Геккель обратил внимание на то, что зародышевые стадии многоклеточных очень сходны с колониальными жгутиковыми и низшими многоклеточными. Так, все многоклеточные проходят стадию однослойного шара — бластулы, которая очень похожа на шарообразную колонию вольвокс. Следующая стадия — двухслойный зародыш, или гаструла, — соответствует строению низших многоклеточных — кишечнополостных. Переход однослойной стадии зародыша в
двухслойную происходит чаще всего путем впячивания одной из сторон шара. На основании этого Геккель предположил, что многоклеточные в своем зародышевом развитии повторяют этапы перехода от одноклеточных к многоклеточным.
По мнению Геккеля, первые многоклеточные возникли следующим образом. У шарообразной колонии типа вольвокс произошло впячивание одной из сторон шара, и образовался простейший двухслойный организм. На месте впячивания формируется рот, внутренний слой выстилает кишку. После того как образовалось два слоя, функции клеток разделились. Наружный слой стал выполнять защитную функцию, а внутренний — пищеварительную. Этот первичный многоклеточный организм Геккель назвал гастреей. Он правильно связал происхождение многоклеточных с колониальными простейшими, но не мог объяснить причин, которые привели к образованию двухслойности.
Теория паренхимеллы. С точки зрения И. И. Мечникова, в однослойных шаровидных колониях второй — внутренний — слой образовался не путем впячивания, а путем выселения клеток из наружного слоя внутрь (иммиграция). Отдельные простейшие однослойной колонии после захвата пищи погружались внутрь шара и образовывали там рыхлую массу (паренхиму). Внутри они переваривали пищу, после чего вновь возвращались на поверхность. Постепенно в процессе эволюции часть клеток, переваривающих пищу, утратила способность возвращаться на поверхность и образовала внутренний слой. Сначала они получали пищу через клетки наружного слоя, а затем развилось ротовое отверстие и полость кишки стала способной сообщаться с внешней средой. Как видно, теория Мечникова объясняет образование простейшего двухслойного организма вполне конкретными причинами — захватом и перевариванием пищи. Этот первичный многоклеточный организм Мечников назвал паренхимеллой, или фагоцителлой.
Филогения прокариот
Современные прокариоты охватывают широкий круг организмов, отличающихся по химическому составу и физиологическим функциям. К ним относятся такие разнородные группы организмов, как бактерии и сине-зеленые водоросли.
Бактерии. Те из бактерий, которые не нуждаются для своего питания ни в каких других животных и растениях и существуют за счет химических реакций, происходящих в растворах газообразных или минеральных веществ, называются хемотрофными. Считается, что хемотрофы были первичными организмами на Земле, когда на ней еще не было необходимых условий для жизни растений и животных. Один из наиболее древних остатков живых существ — это так называемые железобактерии железных руд в сланцах архейского отложения, найденных в Северной Америке. В более ранних отложениях не найдено пока никаких следов жизни.
Между тем среди хемотрофных бактерий железобактерии являются далеко не простейшими по своему строению, и возможно, что от появления на Земле первых живых существ и до времени отложения слоев кристаллических архейских сланцев прошел не один миллион лет.
Среди бактерий есть такие, которые произошли, по всей вероятности, путем упрощения более сложных организмов. Так, среди серных бактерий встречаются представители, очень сходные по окраске с имеющими хлорофилл сине-зелеными водорослями, а живущие в крови высших животных бактерии спирохеты могут иметь животное происхождение — от простейших. Образ жизни бактерий-паразитов способствует упрощению организации благодаря своему однообразию и делает сходными организмы, различные по происхождению. Когда развилась растительная и животная жизнь, то легкость питания органическими веществами, естественно, способствовала развитию бактерий, питающихся за счет животных и растений.
Сине-зеленые водоросли — простейшая группа растительных организмов. Наличие двух фотосистем (ФС1 и ФС2) у сине-зе леных водорослей свидетельствует о высоком уровне их эволю ционного развития. Однако качественный состав фотосистем у сине-зеленых водорослей отличается от высших растений и зеленых водорослей. После изучения состава пигментов (хлорофилл А, ФС1, ФС2, фикобиллины) и их свойств у сине-зеленых водорослей и бактерий (бактериохлорофилл, ФС2) стала очевидной безосновательность попыток выводить одинаковое происхождение сине-зеленых водорослей и бактерий, как это было принято. Сходство между ними скорее внешнее. Таким образом, вероятнее всего, что сине-зеленые водоросли представляют собой самостоятельную ветвь от первоначальных прокариотов. Однако поскольку эти водоросли — прокариоты, было предложено назвать их цианобактериями, чтобы как-то отличать от эукариотических водорослей.
Филогения зеленых водорослей
Зеленые водоросли — прогрессивная группа растительных организмов по сравнению с сине-зелеными. Полагают, что зеленые водоросли возникли из прокариот, содержащих хлорофилл А и В.
В отличие от сине-зеленых в клетках зеленых одноклеточных водорослей содержатся вполне сформировавшиеся хлоропласта, или, как их называют у водорослей, хроматофоры. В дальнейшем определяющим фактором в эволюции водорослей явилась эволюция хроматофора. Наличие у многих представителей водорослей осевого хроматофора позволяет обнаружить и изучить родственные связи зеленых водорослей и проследить их историческое развитие.
Общие направления функциональной эволюции зеленых водорослей можно сформулировать в следующих тезисах.
1. Массивный осевой хроматофор является первичным, более примитивным, и лишь впоследствии, в процессе прогрессивной эволюции, из него возник пристенный, функционально более совершенный и целесообразный для фотосинтетической деятельности. Осевой хроматофор, по мнению академика Л. В. Топачевского, мог возникнуть лишь у амебовидных представителей водорослей, так как при расположении в центре клетки он не мешал образованию псевдоподий и не подвергался их влиянию. Таким образом, амебовидные формы следует считать первичными среди зеленых водорослей. Другие типы строения хроматофора — чашевидный, пластинчатый и особенно пристенный — более прогрессивные и преобладают у подавляющего большинства современных зеленых водорослей.
2. Жгутиковые формы водорослей, среди которых наиболее Простыми являются хламидомонады, имеют вторичное происхождение от амебовидных организмов. Основной критерий обоснования этого вывода — сложность жгутиков и относительная простота строения псевдоподий.
3. Из более простых хламидомонад путем утраты подвижности возникли хлорококковые, а из них — нитчатые улотриксовые.
4. Появление многоядерных клеток положило начало развитию кладофоровых.
5. Конъюгаты, включающие как одноклеточные, так и нитчатые формы (спирогира), представляют собой отдельную группу водорослей, берущую начало от первичных амебовидных форм. Поэтому в цикле их размножения отсутствуют жгутиковые формы.
6. Диатомовые водоросли происходят от жгутиковых организмов, окрашенных в желтый цвет и способных отлагать в своих оболочках небольшое количество кремнезема. Диатомовые характеризуются разнообразием клеточных форм, приспособленностью к свободноплавающему образу жизни или к образу жизни организма, прикрепляющегося под водой к скалам или другим твердым предметам. Они не переходят ни в какой высший тип, являясь как бы законченным в своем развитии.
Филогения высших растений
Рассмотрим основные признаки, приобретенные зелеными нитчатыми водорослями при переходе от одноклеточных форм к многоклеточным, а затем к высшим растениям.
1. Сходство зеленых нитчатых водорослей с высшими растениями прежде всего в крупных размерах их тела (таллома). Они широко распространены в тех же биотопах, что и высшие растения. Возникновение крупных макроскопических талломов способствовало захвату жизненного пространства в борьбе за условия существования.
2. Наличие хорошо развитых вакуолей, связанных в основном с многоклеточностью организмов, что наблюдается у нитчатых водорослей и низших растений. Установлено, что объем протоплазмы клеток зеленых нитчатых водорослей изменяется в зависимости от их фотосинтетической активности. На протяжении световой фазы протопласт увеличивается в размерах за счет уменьшения объема вакуоли. Во время темновой фазы уменьшается протопласт, а объем вакуоли увеличивается. Следовательно, центральная вакуоль — это пространственный резерв для интенсивного роста в процессе фотосинтеза.
3. Появление фотопериодизма. Связь жизненных функций со сменой дня и ночи у нитчатых водорослей аналогична высшим растениям.
4. Предками первичных наземных растений могли быть зеленые нитчатые водоросли с многорядными нитями, поэтому легко представить возникновение флоэмы. Ситовидная трубка является аналогом и гомологом нити зеленых нитчатых водорослей и выполняет ту же функцию проводящей системы. Ксилемы у зеленых нитчатых водорослей нет. Однако процесс ее закладки у высших растений свидетельствует о том, что эта ткань возникла не сразу.
5. У зеленых водорослей впервые наблюдаются различные сочетания ядерных фаз. Обычно у них преобладает гаплоидная фаза (гаметофит) над диплоидной (спорофит), но у более сложных нитчатых водорослей уже наблюдается обратное явление. Поэтому с выходом на сушу дальнейшее сокращение гаметофита вплоть до половых клеток явилось одним из важнейших факторов завоевания различных ареалов с болей жесткими условиями обитания, чем в водной среде.
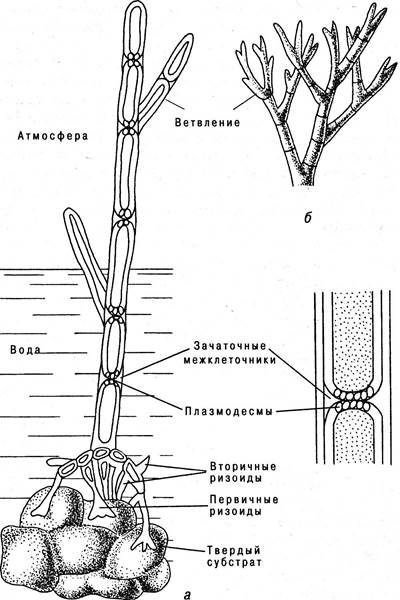
Происхождение высших растений из нитчатых водорослей является наиболее вероятным. Предками этих первичных наземных растений могли быть зеленые нитчатые водоросли с многорядными нитями. Большинство из них прикреплялось к субстрату (камням) с помощью специальных приспособлений — ризоидов, которые являлись продолжением главной оси растения (рис. 6.11).
Рис. 6.11. Зеленая нитчатая водоросль как элементарное высшее растение: а — кладофора; б — хетоморфа
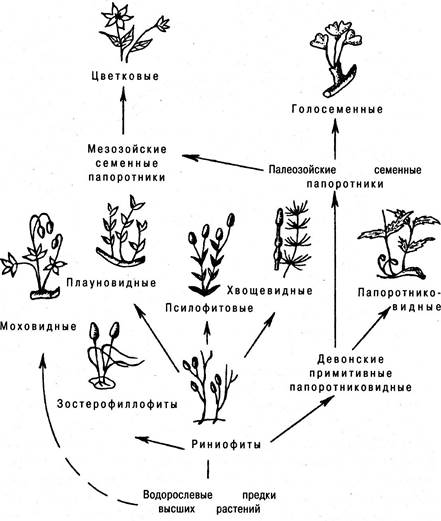
Древнейшие остатки растений обнаружены в раннедевонских отложениях. Эти растения относятся к классу риниопсида (псилофиты). Для представителей этой группы древнейших высших растений характерны дихотомическое (вильчатое) ветвление и отсутствие листьев. Вместо корней у них были ризомоиды, вместо корневых волосков — ризоиды. Корней у риниопсидов еще не было. Они возникли впоследствии из побегов. У таких древнейших растений обнаружена микориза (гриб-симбионт), что нашло отражение в дальнейшей эволюции высших растений. Деление клеток и рост риниопсидов верхушечный. Вторичного роста еще нет. Расположение спорангиев тоже верхушечное. Устьичный аппарат лишь в зачаточном состоянии. Для них характерно чередование гаметофита и спорофита с преобладанием последнего. Именно из риниофитных растений в процессе эволюции возникло большое количество разнообразных наземных форм, в том числе сосудистые растения (рис. 6.12).
Рис. 6.12. Схема эволюции высших растений
Происхождение красных и бурых водорослей. Это своеобразные группы водорослей как по морфологическому строению слоевища, так и по биохимическому составу. Прежде всею следует подчеркнуть, что они — эукариоты. Главным отличием этих водорослей является наличие в них кроме хлорофилла А каротиноидов и дополнительных пигментов: в красных — фикоэритрина и фикоцианина, а в бурых — фукоксантина. Эти типы растений имеют значительную филогенетическую обособленность. У них не прослеживается переход от одноклеточной организации к многоклеточной. Вместе с тем в эволюционной цепи многоклеточные водоросли (бурые и багрянки) зашли в тупик, достигнув значительного расцвета и высокой степени дифференциации в пределах типа растительного царства. Можно лишь только предположить их эволюционное родство с низшими представителями зеленых водорослей.
Происхождение мохообразных. Мохообразные происходят от водорослей, у которых гаметофит преобладает над спорофитом. Гаметофит эволюционировал из простертой части многонитчатого таллома, а спорофит — из вертикальной. Данная группа растений стоит особняком в филогенетическом отношении. Это слепая ветвь эволюции. Возникновение мохообразных происходило в силуре.
Происхождение плауновидных. Среди современных высших растений плауновидные — одна из древнейших групп. Они возникли в середине девона, отчленившись от растений, близких к риниофитам, которые имели антеридии с двухжгутиковыми сперматозоидами. Процветали плауновидные в палеозое, когда существовали их гигантские древовидные формы (лепидодендроны).
Происхождение хвощевидных. Появились хвощевидные в девоне и процветали от карбона до триаса. Несмотря на членистость стеблевых органов и колосков, они по многим признакам напоминают риниофиты. Вероятно, древние хвощи произошли от растений, близких к риниофитам, имевших многожгутиковые сперматозоиды в антеридиях. Древовидные хвощи (каламиты) наряду с древними плауновидными образовали леса каменноугольного периода.
Происхождение папоротникообразных. Древнейшие ископаемые формы известны с девона. В карбоне крупные древовидные папоротники наравне с другими споровыми растениями составляли обширные влажные леса, остатки которых участвовали в образовании залежей каменного угля. Наряду с возникновением типичной сосудистой системы у папоротников появились также листья и корни. По своему происхождению (от древних риниофитов) папоротники имеют значительно больше родства с хвощевидными, чем с плауновидными.
Происхождение голосеменных. Первыми голосеменными растениями стали семенные папоротники, которые размножались уже не спорами, а семенами. Важнейшая особенность семени — наличие в нем зародыша, защищенного от неблагоприятных условий среды и мутагенных факторов, а также запасных веществ — белков, углеводов и жиров, необходимых для роста зародыша. Оплодотворение у семенных папоротников происходило уже не в водной среде. Это дало им возможность избежать массового вымирания сначала в пермском
периоде, а затем в триасовом периоде, когда климат стал более сухим. Таким образом, расцвет голосеменных относится к концу палеозоя и мезозою — эпохе горообразования, поднятия материков и проявления магматизма.
Происхождение покрытосеменных. В середине мелового пери ода (около 120 млн лет тому назад) по не вполне понятным причинам наступила вспышка видообразования цветковых растении, в результате чего к концу данного периода покрытосеменные за воевали сушу. Господство покрытосеменных продолжается в течение всей кайнозойской эры до настоящего времени. Важнейшая особенность покрытосеменных растений — возникновение цветка, открывшего возможность перекрестного оплодотворения путем переноса пыльцы сначала ветром, а затем и насекомыми. Цветок в процессе эволюции возник из сильно укороченного листостебельного побега. Чашелистики, лепестки, тычинки и пестик — это видоизмененные листья, которые у предков цветковых растений окружали органы размножения. Крупным эволюционным достижением цветковых растений является двойное оплодотворение, открытое в 1989 г. С. Г. Навашиным. Вероятным предком покрытосеменных считают семенные папоротники, обитавшие в мезозойской эре.
Филогения животных
Наиболее известна науке история животных. Существует точка зрения, что губки возникли от какого-то одного предка, принадлежащего простейшим, а остальные многоклеточные животные — от какого-то другого. Самые крупные категории, на которые делятся животные, — это типы; на протяжении геологического времени возникло по меньшей мере 35 типов, из которых 26 существуют до сих пор, а 9 вымерли (рис. 6.13).
Рис. 6.13. Ныне живущие типы животных (сгруппированы в соответствии с планом их строения; для каждого типа приведено изображение одного из его представителей)

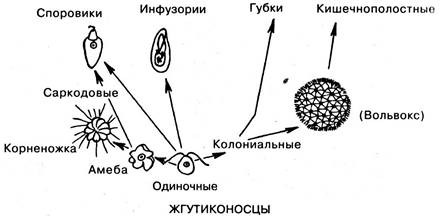
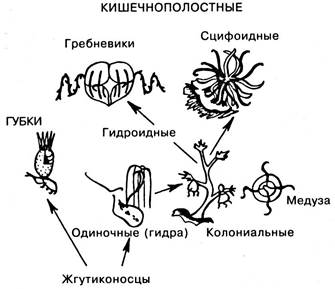
Происхождение простейших. Тип простейших произошел от жгутиконосцев, давшим начало голым, а затем раковинным корненожкам, а также, возможно, инфузориям. Что касается споровиков, то часть их происходит от амеб, а другие формы этого класса непосредственно от жгутиконосцев. Одиночные жгутиковые дают начало колониальным формам, причем потомками шарообразных колоний жгутиковых являются кишечнополостные, а из других колониальных форм, жгуты которых окружены плазматическими воротничками, развились губки (рис. 6.14).
Рис. 6.14. Филогения простейших
В настоящее время традиционной точке зрения о происхождении инфузорий от жгутиконосцев можно противопоставить представление о возникновении инфузорий от многоклеточных плоских червей. В ходе развития клеточные границы исчезли, а соматические ядра слились в единый макронуклеус, обладающий сверхвысокой плоидностью. Гипотеза о происхождении инфузорий от многоклеточных животных, утерявших клеточные границы, была высказана в 1924 г. И. И. Месяцевым.
В пользу данной гипотезы можно привести следующие аргументы.
1. Имеется некоторая аналогия между половым процессом у инфузорий и плоских червей. Спаривание плоских червей внешне напоминает конъюгацию инфузорий.
2. У мелких форм турбеллярий, как и у инфузорий, движение осуществляется преимущественно за счет ресничек. У инфузорий неоднократно обнаруживали многоклеточность, а также нервные узлы и нервные пучки, что является особенностью многоклеточных организмов.
Происхождение губок и кишечнополостных (рис. 6.15). Как уже сказано, губки и одиночные гидроиды происходят от жгутиконосцев. Одиночные гидроиды дают начало колониальным формам, а от последних развиваются сцифоидные и гребневики. Гребневики образуют переход к ресничным червям, т. е. к трехслойным животным. Об этом свидетельствует появление впервые у гребневиков следующих прогрессивных признаков.
Рис. 6.15. Филогения губок и кишечнополостных
1. Развитие между экто- и энтодермой зачатка третьего зародышевого листка — мезодермы. За счет этого зачатка у взрослых форм в студенистом веществе мезоглии развиваются многочисленные мышечные элементы.
2. В связи с переходом от плавания к ползанию впервые обособились вентральная и дорзальная стороны тела.
3. Впервые появляются элементы двусторонней (билатеральной) симметрии.
4. Нервные клетки начинают концентрироваться в нервные узлы (ганглии).
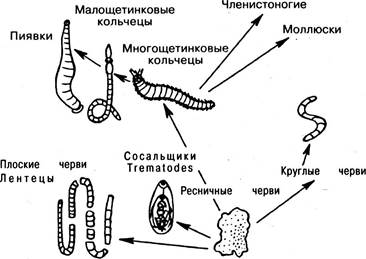
Происхождение червей (рис. 6.16). Ресничные черви дают начало плоским (сосальщикам и ленточным), круглым и кольчатым червям. Из последних многощетинковые черви имеют наиболее примитивное устройство, и от них происходят более поздние и специализированные формы (многощетинковые и пиявки). От многощетинковых кольчецов также обособились два новых типа животных — членистоногие и моллюски.
Рис. 6.16. Филогения червей
Происхождение моллюсков. Ближе всех к кольчецам класс боконервных моллюсков; более высокодифференцированные — брюхоногие и пластинчатожаберные; высший класс составляют головоногие (осьминоги и каракатицы).
Происхождение членистоногих (рис. 6.17). Весьма специи лизированный тип членистоногих, происходя от кольчатых червей, очень давно разделился на две ветви. Одна из них имеет исходной формой ископаемых — трилобитов, от которых произошли, с одной стороны, ракообразные, с другой ракоскорпионы, давшие начало паукообразным. Другая ветвь ведет к насекомым через древнюю форму первичнотрахейных, отдельные представители которых сохранились до нашего времени, и через филогенетически более поздних многоножек (рис. 6.18).
Рис. 6.17. Филогения членистоногих
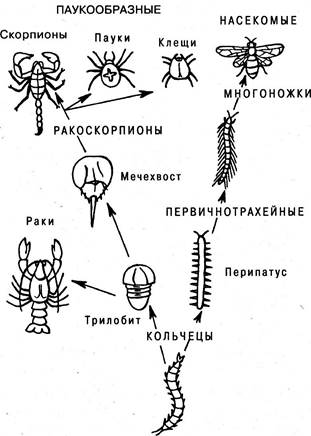
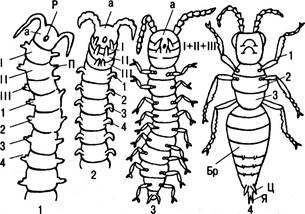
Рис. 6.18. Происхождение насекомых (схематично): 1 — кольчатые черви; П — параподии; Р — рот; а — головной конец; 2 — смещение 1, 2, 3 сегментов-члеников кпереди, сегменты 1+2+3 образовали сросшуюся воедино голову. Их придатки стали челюстями; 4 — насекомые; позади головы обособляются 1, 2 и 3 членики, образуя грудь. Остальные образовали брюшко (Бр). На нем остатки конечностей (Ц) и яйцеклад (Я), иногда становящийся жалом
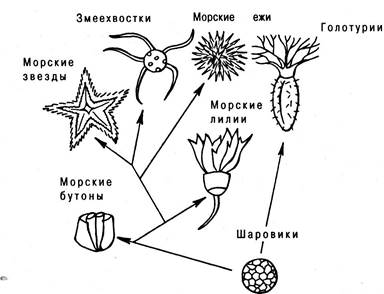
Происхождение иглокожих (рис. 6.19). Предки иглокожих были двусторонне симметричными животными, перешедши ми потом к сидячему образу жизни; к этому времени (начало палеозойской эры) относится и возникновение лучевой симметрии. Современные иглокожие в большинстве случаев ведут свободный образ жизни (кроме морских лилий) и сохраняют лучевую симметрию (кроме сердцевидных морских ежей и голотурий, у которых наблюдается частичный возврат к двусторонней симметрии). Наиболее древними из известных предков иглокожих являются шаровики (начало палеозоя); от них возникли морские бутоны (каменноугольный период), а от близких к последним форм пошло постепенное усложнение организации через морских лилий к морским звездам, офиурам и морским ежам. Голотурии отделились от общего ствола иглокожих очень рано, происходя непосредственно от шаровиков.
Рис. 6.19. Филогения иглокожих
Филогения хордовых
Происхождение первых позвоночных (рис. 6.20). Предками позвоночных были наиболее просто организованные беспозвоночные из всех существовавших в прошлое время. Это простые, сидячие формы, напоминающие ныне существующих глубоководных морских животных, относящихся к классу крыложаберных. Хотя крыложаберных относят к типу хордовых, тело этих животных, прикрепленное ко дну с помощью стебелька, в основном состоит из пищеварительного тракта. Вокруг рта находятся щупальца, служащие для захвата пищевых частиц, приносимых водой. Дальнейшее развитие этих простейших существ, по-видимому, завершилось появлением жаберных щелей, которые представляют собой парные отверстия, открывающиеся по бокам тела и ведущие из глотки наружу.
Рис. 6.20. Схема, показывающая возникновение первых позвоночных из личиночной формы животных, напоминающих оболочников, в результате педоморфоза
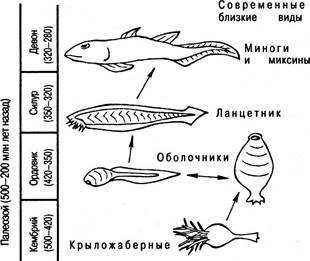
Первоначально жаберные щели служили фильтрами для задержания пищевых частиц, в дальнейшем же они превратились в аппарат для дыхания. На этой стадии эволюционного процесса наши предки, должно быть, напоминали современных оболочников. Оболочники, как сидячие, питающиеся за счет фильтрования воды формы, представляют собой эволюционный тупик. Что же касается их личинок, то в процессе эволюции неоднократно имело место явление так называемого педоморфоза, когда взрослые особи элиминировались, а личиночные формы становились половозрелыми и способными к воспроизведению себе подобных. Вполне вероятно, что в те древнейшие времена личинки животных, напоминавших оболочников, имели жаберный аппарат на уровне глотки; мускульный хвост, усиленный толстой гибкой продольной струной; дорзальный нервный тяж, который в головном отделе получал сенсорно информацию от рудиментарных органов чувств. Первые хордовые, возникшие путем педоморфоза, по всей вероятности, мало отличались от современных ланцетников. Ископаемые остатки небольших примитивных хордовых, не имевших щупалец и челюстей, обнаружены в силурских отложениях. В девонском периоде уже в изобилии появляются первые позвоночные, представленные рыбообразными бесчелюстными животными.
Рыбы, давшие начало наземным позвоночным. Первым большим достижением ранних позвоночных было развитие челюстей. Однако и до наших дней дошли такие реликты, как круглоротые (миноги, миксины). Основные пути эволюции рыб сопровождались усовершенствованием челюстей с современным развитием жаберных крышек, защищающих жабры, и окостенением скелета. Так появился класс костистых рыб.
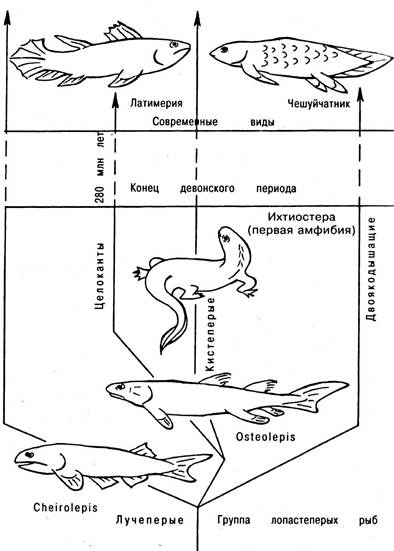
В середине девона костные рыбы дивергируют в двух различных направлениях, различаясь главным образом строением плавников. У одних плавники не имели покрытых чешуей лопастей основания, у других основания плавником напоминали мясистую лопасть. У последних передняя пари плавников, расположенных на груди, могла легко трансформироваться в переднюю пару конечностей наземных. Задняя пара плавников, расположенная позади средней части туловища, также могла превратиться в заднюю пару конечностей. Совершенно очевидно, что только примитивные костные рыбы девонского периода, обитавшие в мелких, периодически пересыхавших внутриматериковых водоемах, были существами достаточно “земноводными” для того, чтобы дать начало наземным позвоночным (рис. 6.21). В классе рыб представители лопастеперых оказались, однако, не особенно преуспевающими. В настоящее время только три рода двоякодышащих рыб, которые до сих пор обитают и мелких пресных водоемах, и один вид латимерии, живущих в океане, являются представителями этой эволюционной ветви (рис. 6.22).
Рис. 6.21. Возникновение и развитие позвоночных (снизу-вверх): а — кистеперые рыбы, выходящие на сушу; б — первые земноводные; в — первые пресмыкающиеся, уже откладывающие яйца на суше

Рис. 6.22. Деление девонских рыб на лучеперых и лопастеперых
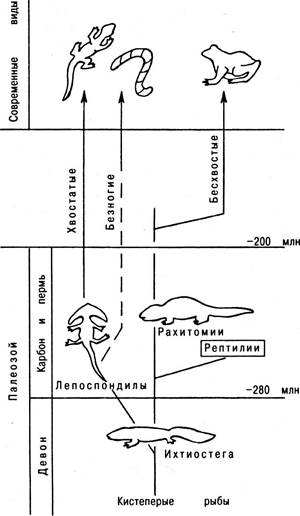
Происхождение амфибий. Первыми амфибиями, ведущими свое происхождение от вида, близкого к костным рыбам, были ихтиостеги. У них имелись уже хорошо развитые плечевой и тазовый пояса, с помощью которых они управляли расчлененными передними и задними конечностями. Такие конечности вполне могли обеспечить передвижение животного по суше. Развитие амфибий в каменноугольном и пермском периодах было следующим.
1. Развитие прочной кожи, часто покрытой костными пластинами, предохраняющими от потери влаги в результате испарения.
2. Укрепление позвоночного столба вследствие формирования цельных позвонков, которые с помощью мышц и связок образуют прочный горизонтальный ствол, поддерживающий тело.
3. Усовершенствование дыхательной системы путем образования хорошо развитых носовых ходов, которые открываются ноздрями, расположенными на дорзальной поверхности черепа.
Одни из самых первых потомков ихтиостег, известных под общим названием Эмболомеры, уже в раннем карбоне определились в направлении развития как будущие рептилии. Тем временем основная ветвь амфибий (так называемые Лабиринтодонты, среди них Рахитомии) развивались в направлении современных, ныне живущих лягушек и жаб. Предки же хвостатых земноводных (тритонов, саламандр, протеев) оказались совсем иного происхождения. С самого начала эволюции амфибий существовала небольшая ветвь, которая резко отличалась от основной ветви, получившей общее название Лепоспондилы (рис. 6.23). Современные хвостатые амфибии как раз и представляют собой потомков древней группы Лепоспондилов.
Рис. 6.23. Филогения амфибий
Появление высших позвоночных — рептилий и птиц. Во время каменноугольного периода произошло крупнейшее событие в эволюции наземных позвоночных — животные приобрели ряд приспособлений к развитию яйца вне водной среды. Первыми животными, у которых выработался ряд таких приспособлений, были рептилии. Так, благодаря появлению яиц с зародышевыми оболочками отпала необходимость в личиночной стадии в процессе онтогенетического развития. Все зародышевые структуры заключены в скорлупу, достаточно прочную для того, чтобы защищать содержимое яйца, и в то же время достаточно пористую для того, чтобы обеспечить доступ кислорода в яйцо и выделение из него углекислого газа.
Превращение одной из наиболее древних лабиринтообразных амфибий (Эмболомеры) в рептилий (Котилозавров) произошло в конце пермского периода. На первом этапе дивергенции потомки Котилозавров образовали пять подклассов, три из которых нас интересуют особенно: Диапсида, Анапсида и Синапсида. Из этих трех подклассов Диапсида была преуспевающей ветвью рептилий (рис. 6.24). Они характеризовались наличием двух задних височных ям, разделенных заглазничной и чешуйчатой костями.
Рис. 6.24. Три подкласса рептилий и их потомки
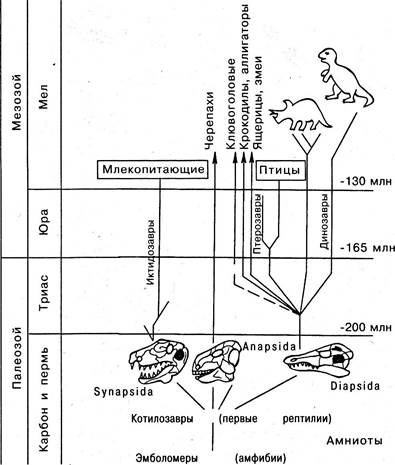
В начале мезозойской эры (триас) ветвь подкласса Диапсид представлена была так называемыми архозаврами. История эволюции этой ветви рептилий, кульминацией которой было появление динозавров, достаточно хорошо известна. Рептилии, принадлежащие к подклассам Анапсида и Синапсида, занимали весьма скромное положение. Кроме динозавров архозавры дали также начало новому классу позвоночных — птицам. Как класс птицы произошли от могущественной ветви
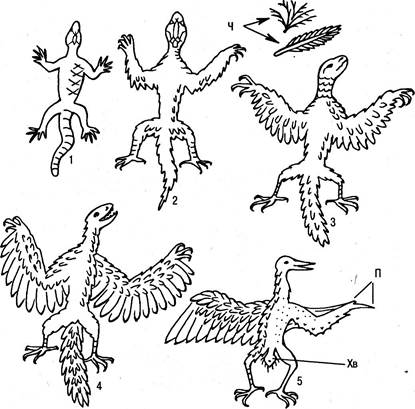
архозавров, когда последние находились в зените своего расцвета. Может показаться, что данный случай противоречит общему правилу, согласно которому только примитивные виды какого-либо класса делают большой скачок и дают начало новому, более совершенному классу. Следует сказать, однако, что процесс возникновения птиц из рептилий не сопровождался появлением каких-либо крупных новшеств. Поэтому не удивительно, что птиц часто называют пернатыми рептилиями (рис. 6.25).
Рис. 6.25. Происхождение птиц. Чешуя (Ч) превращается в перья и пух, сохраняясь на ногах: 1 — древние ящерицы; 2 — 3 — лазающие формы; 4 — археоптерикс; 5 — птицы (правая сторона без перьев, чтобы показать остатки пальцев (П) и укоротившийся хвост (Хв)
Ныне живущие ящерицы и змеи, а также крокодилы и аллигаторы произошли от рептилий, относящихся к подклассу Диапсида. Напротив, черепахи являются потомками Анапсида. В отличие от Диапсида у Анапсида отсутствуют височные и мы позади глаз.
Появление млекопитающих. Представители подкласса Синапсида так и не достигли господствующего положения как рептилии. Непосредственно продолжая линию примитивных рептилий, эта группа дала начало первым млекопитающим. Именно они являются соединяющим звеном примитивных рептилий, с одной стороны, и млекопитающих — с другой (рис. 6.26).
Рис. 6.26. Происхождение млекопитающих (1 — 4) от звероящеров до первых яйцекладущих млекопитающих: В — расположение волос, растущих между чешуйками. Видно, как чешуя постепенно заменялась волосяным покровом; М — расположение млечных желез
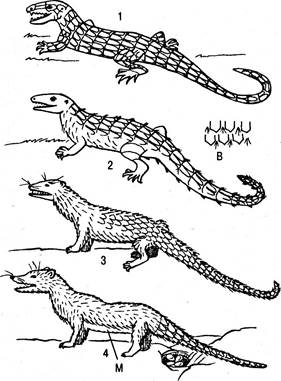
Характерной чертой подкласса было наличие одной височной ямы, ограниченной заглазничной и чешуйчатой костями. В отличие от Диапсида представители Синапсида на протяжении всей своей эволюции сохранили горизонтальное положение тела и передвигались с помощью четырех конечностей. Среди Синапсид наиболее близким к млекопитающим были представители, известные под общим названием Териодонты. Позднее появилась группа рептилий, которая служила промежуточным звеном между прогрессивными Териодонтами и примитивными млекопитающими. К таким рептилиям относились Ихтиозавры. Несмотря на то что эта группа рептилий как бы заложила фундамент для последующего развития и триумфа млекопитающих уже в раннем мезозое, прошли многие миллионы лет, прежде чем наступил век млекопитающих.
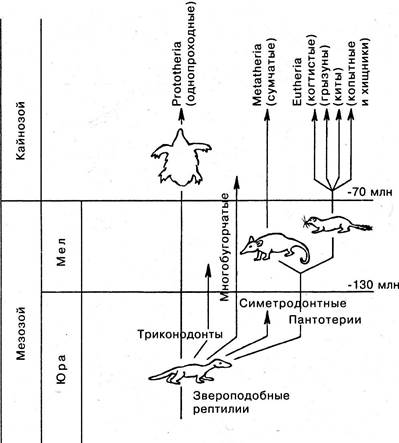
Ныне живущие млекопитающие принадлежат к трем подклассам: Однопроходные (Яйцекладущие, или Клоачные), Сумчатые и Плацентарные (рис. 6.27).
Рис. 6.27. Схема, иллюстрирующая происхождение однопроходных, сумчатых и плацентарных млекопитающих от звероподобных рептилий
Происхождение приматов (рис. 6.28). Самыми первыми и наиболее примитивными плацентарными млекопитающими были насекомоядные, и, очевидно, именно они были родоначальниками всех других плацентарных. От этой ветви примитивных насекомоядных произошло 28 отрядов плацентарных млекопитающих, 11 из которых к настоящему времени вымерли.
Рис. 6.28. Схема родословного древа приматов
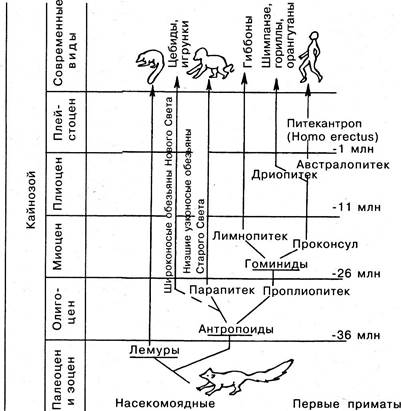
От какого-то гипотетического насекомоядного предка, обитавшего на деревьях и напоминавшего тупайю, приматы развивались в трех главных направлениях. Первая радиация примитивных приматов в палеоцене (50 — 70 млн лет назад) привела к возникновению подотряда, к которому относятся лемуры, лори и долгопяты.
В результате второй радиации в раннем эоцене (примерно 40 млн лет назад) появился другой подотряд. К нему принадлежат все обезьяны Старого и Нового Света. Третья радиация м раннем олигоцене (приблизительно 35 млн лет назад) привела к образованию надсемейства Гоминид. Предок Гоминид проплиопитек обладал тенденцией к увеличению тела и мозга. Следом за проплиопитеком появились лимнопитеки и проконсул. Лимнопитеки, по-видимому, представляли собой боковую ветвь, которая дала начало современным гиббонам, тогда как менее специализированный проконсул стал предком современных Гоминид. Потомками проконсула были различные дриопитеки. Эти обезьяны, вероятно, напоминавшие по внешнему виду шимпанзе, были предками не только современных шимпанзе, горилл и орангутанов, но, возможно, и нас самих.
Антропогенез
Современная антропология выделяет несколько этапов антропогенеза.
I этап — предшественники человека. В основании развития линии приматов, приведшей к возникновению Человека, находятся австралопитеки (южная обезьяна). Находки австралопитеков сделаны на территории главным образом Южной Африки.
Австралопитеки — достаточно крупные обезьяны (масса приблизительно 20 — 50 кг, рост 120 — 150 см), ходили на двух ногах при выпрямленном положении тела. Масса мозга была близка к 550 г. Зубы расположены в виде широкой, как у человека, а не узкой, как у обезьяны, дуги. Клыки у них были небольшие. Австралопитеки — обитатели открытых пространств, жившие в пещерах. Они широко использовали различные кости животных, в качестве ударных орудий применяли палки, камни. Австралопитеки вымерли до окончания плейстоцена (т. е. они жили в период от 9 млн до 750 000 лет тому назад). Среди прогрессивных австралопитеков выделяется Человек умелый (Homo habilis). Масса мозга у него была больше, чем у среднего австралопитека (650 г). Первый палец стопы Человека умелого не был отведен в сторону, как у обезьяны; концевые кисти фаланг были короткими и плоскими, как у настоящего человеческого существа. Человек умелый уже мог изготовлять грубые галечные орудия. Это произошло 2,6 млн лет тому назад. Значение находки галечных орудий огромно, так как трудовая деятельность является границей, отделяющей Человека от остальной природы.
II этап — древнейшие люди (архантропы). Внешне архантропы были уже весьма похожи на современного человека. Масса
мозга у них составляла около 800 — 1000 г. Именно при такой массе мозга становится возможным овладение речью. У архантропов прогрессивно развивалась способность к производству орудий труда. Средний рост мужчин был около 160 см, а женщин — 150 см. С зоологической точки зрения эти формы, видимо, относятся к одному большому виду — Человеку выпрямленному (Homo erectus).
Занимая промежуточное положение между современным человеком, с одной стороны, и австралопитеком, с другой стороны, архантропы начали быстро расселяться по Африке, Средиземноморью, Южной, Центральной и Юго-Восточной Азии. Вероятно, в этот же период начал использоваться огонь. Расселяясь и попадая в новые условия существования, архантропы образовали отдельные изолированные формы. Сейчас изучено довольно значительное число форм древнейших людей. Наиболее известные: питекантроп (о. Ява), синантроп (Китай), гейдельбергский человек (Средняя Европа), атлантроп (Алжир), телантроп (Южная Африка), олдовайский питекантроп (Центральная Африка).
III этап — древние люди (палеонтропы). Древние люди занимают промежуточное положение между древнейшими людьми — архантропами и ископаемыми формами Человека разумного — как по строению тела, так и по развитию культуры. По месту первого описания таких людей (долина Неандерталь близ Дюссельдорфа) их часто называют неандертальцами. Неандертальцы имели сравнительно небольшой рост (155 — 165 см). Масса их мозга составляла около 1500 г., при чем сильное развитие получили отделы, связанные с логическим мышлением. Орудия неандертальцев — ручные рубила, остроконечники и скребки — были более совершенными, чем примитивные обработанные гальки.
Увеличение числа находок позволило выяснить, что неандертальцы были весьма неоднородной группой. Они подразделялись на две главные расы: поздние неандертальцы и ранние неандертальцы. Поздние неандертальцы выглядел и звероподобными, однако масса их мозга превышала массу мозга современного человека (1600 г). Среди них были развиты внутригрупповые связи и на охоте, и при защите от врагов, и от неблагоприятных погодных условий. Представители поздних неандертальцев очень небольшими группами могли выживать и побеждать в борьбе за существование. Ранние неандертальцы оказались на совершенно ином эволюционном пути — они шли к победе в борьбе за существование через объединение сил отдельных индивидов, что привело 50 — 40 тыс. лет назад к возникновению того вида, к которому принадлежим и мы с вами, — Человека разумного (Homo sapiens).
IV этап — современные люди (неантропы). Такие качества неантропов (кроманьонцев), как развитие речи, интеллект, коллективная трудовая деятельность, привели в процессе дальнейшего развития к возникновению человеческого общества.
Существуют две основные точки зрения на происхождение человека современного типа. Первая точка зрения — гипотеза полицентризма, вторая — гипотеза моноцентризма.Согласно первой гипотезе Человек разумный возник в нескольких местах планеты из разных предковых форм, принадлежащих к палеонтропам (или даже к архантропам), согласно второй — существовало единое место возникновения человечества из какого-то общего предкового ствола. В последнее время получила всестороннюю аргументацию гипотеза широкого моноцентризма Я. Я. Рогинского. Согласно этой гипотезе человек возник в одном месте, после чего началось широкое и активное расселение возникших неантропов по планете — они быстро захватили Переднюю Азию, Европу, Центральную и Южную Африку.
Дифференциация Человека разумного на расы. Все современное человечество принадлежит к единому полиморфному виду — Человек разумный. Единство человечества основано на общности происхождения, социально-психического развития, на неограниченной способности к скрещиванию людей различных рас, а также на практически одинаковом уровне общего физического и умственного развития представителей всех рас.
Вид Человек разумный распадается на три большие расы — европеоидную, монголоидную и негроидную. В пределах каждой большой расы выделяют еще малые, или вторичные, расы. Между большими расами существуют переходные (рис. 6.29). Механизм образования рас человека таков же, как и механизм возникновения внутривидовых группировок у животных.
Рис. 6.29. Схема взаимоотношения современных рас человека
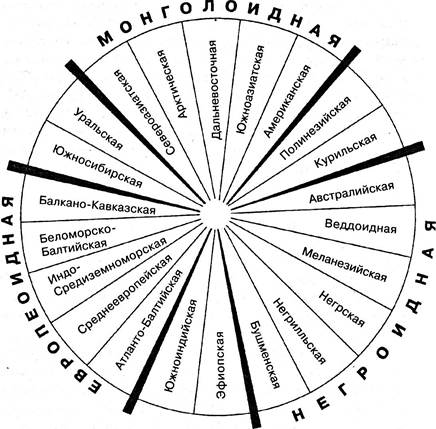
Происхождение и развитие рас представляет собой очень сложный исторический процесс. Около 100 тыс. лет назад на Азиатском континенте с прилегающими к нему областями Африки и Европы сформировались две расовые группы — юго- западная и северо-восточная.
Эти группы разделялись горными цепями Гималаев и Гиндукуша и хребтом Индокитая. Первая из этих ветвей позднее дала начало европеоидной и негроидной большим расам. Вторая же сформировала монголоидную расу, распавшуюся на ряд мелких ветвей, представители одной из которых, американской, проникли в районе Берингова пролива из Азии в Северную Америку и затем заселили весь Новый Свет.
Различия между расами касаются лишь второстепенных признаков, обычно связанных с частными приспособлениями к конкретным условиям существования. Сторонники расизма в своем утверждении о биологической неравноценности рас исходят главным образом из происхождения их от разных предков. Они считают, что европеоидная раса ведет свое начало от кроманьонцев, тогда как все остальные связаны и своем происхождении с неандертальцами. Однако эта концепция основана на извращении научных данных. Палеонтология все больше и больше накапливает факты, свидетельствующие о том, что прародина современного человека занимала обширную территорию, но без резко обозначенных изолированных центров формирования рас, как это утверждает теория полицентрического расообразования. Труды ученых подтверждают, что расовые особенности не играют роли в развитии психической деятельности человека. Далее история учит, что человеческая культура не есть результат умственного развития какой-нибудь одной расы, она возникла в процессе многовековой трудовой деятельности всего человечества.
Факторы антропогенеза
1. Прямохождение. Предпосылкой для превращения предчеловека в человека было такое изменение образа его жизни, при котором он начал отвыкать от помощи рук при ходьбе, усваивая прямую походку. Рука стала свободной и могла теперь приобретать новые навыки.
2. Труд. Первое основное условие всей человеческой жизни. Человек имеет орган труда — руку; у животных, в том числе обезьян, их конечности служат лишь органами передвижения. И процессе трудовой деятельности человек производит все необходимое для своей жизни. Начало трудовой деятельности, связанное с использованием передних конечностей, относится к периоду 3 — 2,5 млн лет тому назад.
3. Открытие и использование огня. Это событие привело к активному потреблению мясной пищи, без которой человек не мог стать человеком. Дело в том, что человеку, как и другим млекопитающим, нужны незаменимые аминокислоты, синтезировать которые его организм не может (например, метионин, триптофан). Содержание незаменимых аминокислот в растительной пище далеко не оптимально. Чтобы получить необходимую норму, нужно съесть очень много растений. Жизнь в этом случае превращается в непрерывное поедание, ни на что другое времени не остается.
4. Стадный образ жизни. Способность выполнять несколько важных функций. Члены, стада объединяются для удержания территории и защиты от хищников. Кроме того, что очень важно, стадо создает ту среду, в которой происходит передача опыта друг другу и обучение детенышей другого рода жизненно важным навыкам.
5. Возникновение речи. Потребность в передаче информации и координации деятельности отдельных особей в стадах привело к развитию языка. Обучение языкам требует не только наличия природных интеллектуальных особенностей, но и времени (особенно времени периода развития, предшествующего половой зрелости).
Человекообразные обезьяны, возможно, обладают способностями, необходимыми для того, чтобы выучиться основам человеческого языка, но не могут реализовать эти способности, так как слишком быстро достигают зрелости.
6. Развитие высшей нервной деятельности. Трудовая деятельность и речь вместе со стадным образом жизни за последние несколько сотен тысяч лет привели человека к наращиванию массы головного мозга. Развитие мозга повысило способность к обучению и в свою очередь оказало влияние на трудовую деятельность людей.
7. Развитие альтруистических наклонностей. Трудовая деятельность человека требовала передачи накопленного опыта своим современникам, объединяя усилия отдельных индивидов в процессе труда, охоты и защиты. Происходила грандиозная переоценка ценностей.
Преимущества перед другими получили те племена, которые “догадались” позаботиться о стариках и поддерживать физически не таких сильных, но полезных своими умственными способностями особей (мастера по выделке орудий, хорошие следопыты и наблюдатели и т. п.). Так возникла сложная структура стада, племени, орды. Так возникло Человечество, т. е. Человек вышел за рамки биологических законов, сделав возможным освоение окружающей среды (приручение животных и окультуривание растений), а также создание искусства.
Культурная эволюция
Эволюция человека состоит из двух процессов: органической эволюции и культурной эволюции. Направление органической эволюции приводится в движение длительным отбором. Культурная эволюция имеет собственные движущие силы, но, однако, в рамках естественного отбора. В эпоху культурной эволюции естественный отбор продолжает действовать, и органическая эволюция не прекращается, только в конкурентную борьбу (отбор) вступают такие признаки, как интеллект, способность к обучаемости, восприятию культуры и т. п.
К факторам культурной эволюции относятся:
а) общее накопление культурного наследия;
б) тенденции в развитии культуры, возникающие в результате конкуренции между сообществами, различающимися в культурном отношении (но не генетически).
Принято считать, что культурная эволюция совершается путем постепенного накопления культурного наследия. Сначала происходит приобретение полезных знаний, затем эти знания включаются в общую массу знаний древнего общества, после чего передаются последующим поколениям путем обучения. Каждое следующее поколение добавляет к этому запасу знаний то, что ему удается открыть самому. Новые знания передаются дальше тем же путем. Следовательно, культурное наследие имеет кумулятивный характер.